Synonyms: Gnorimoschema absoluta (Meyrick, 1917) Clarke, 1962![]()
Phthorimaea absoluta (Meyrick, 1917)
Scrobipalpula absoluta (Meyrick, 1917) Povolny, 1964
Scrobipalpuloides absoluta (Meyrick, 1917) Povolny, 1987
Taxonomic position: Insecta, Lepidoptera, Gelechiidae
Authors: N. Mujica, P. Carhuapoma, & J. Kroschel
Common names
Tomato leaf miner, tomato borer, South American tomato moth, South American tomato pinworm (English); cogollero del tomate, gusano minador del tomate, oruga minadora de hoja y tallo, perforador de las hojas del tomate; polilla del tomate, gusano minador de la hoja y del tubérculo de la papa (Spanish); traça-do-tomateiro (Portuguese); mineuse de la tomate (French).
Hosts
Tuta absoluta attacks species of cultivated Solanaceae as Solanum lycopersicum L. (tomato) and Solanum tuberosum L. (potato), Solanum melongena L. (eggplant), Solanum muricatum L. (sweet cucumber), and, Nicotiana tabacum L. (tobacco). Other cultivated host plants are Amaranthus viridis (slender amaranth), Physalis angulata (cutleaf groundcherry), Solanum woronowii, Sorghum halepense (Johnson grass), Xanthium strumarium (common cocklebur). Additionally the pest has also been found to attack other non-cultivated Solanaceae such as Lycopersicon hirsutum, Lycopersicum puberulum Ph, Solanum americanum, S. bonariense L., S. elaeagnifolium L., S. hirtum, S. lyratum, S. nigrum L., S. puberulum, S. sisymbriifolium Lam., S. saponaceum, Physalis angulata, Datura stramonium (jimsonweed), D. ferox and Nicotiana glauca Graham, etc. Lycium chilense (Bertero), Lycopersicon pennellii var. puberulum (Correll), Solanum dubium, Solanum puberulum Phil. Since becoming established in Europe, T. absoluta has also occasionally been found on several species of plants, such as the Cape gooseberry (Physalis peruviana), green beans (Phaseolus vulgaris), Lycium sp. and Malva sp. (Malvaceae). This indicates that T. absoluta shows a high propensity to use various plants as secondary hosts, notably species within the Solanaceae.
Detection and identification
Tomato plants can be attacked from seedlings to mature plants. In tomato infestation found on apical buds, leaves (Photo 1A), and stems (Photo 1B), flowers and fruits (Photo 1AC, D), on which the black frass is visible. Neonate larvae penetrate into tomato leaves, fruits or stems, creating conspicuous mines and galleries. On leaves, larvae feed only on mesophyll tissues, leaving the epidermis intact. Leaf mines are irregular and may later become necrotic. Galleries in stems alter the general development of the plants. Fruits can be attacked as soon as they are formed, and the galleries bored inside them can be invaded by secondary pathogens leading to fruit rot. On potato, T. absoluta attacks aerial parts and occasionally tubers. Foliar damage is as a result of mine-formation within the mesophyll by the feeding larvae (Photo 1E). The newly emerged larvae are quite mobile, and begin their feeding by building galleries on the leaflets. Depending on the place of oviposition and emergence of the larva, it may penetrate the leaf mesophyll, leaf petiole and young shoots. The mines expand as the larva develops causing deformation or rot of the same. The larvae can migrate to another part of the plant, especially when they reach their maximum development or when the affected foliage withers and dries.

Photos 1. Symptoms of the tomato leafminer, Tuta absoluta: in (A) leaves,(B) steam and (C, D) fruits of tomato and field damage and external tuber damage in potato. Photos: Courtesy of CIP.
Morphology
Egg
Eggs are oval in shape, on average 0.38 mm long by 0.21 wide. Newly-laid eggs are creamy white; when mature turn dark and the outline of the larval head capsule can be seen through the chorion; this is called the blackhead stage (Photo 2A).
Larva
First-instar larvae are colorless after hatching, turning to pale yellow-orange. Later instars are yellow-orange and the third-instar larvae measure 3.2 x 1 mm (Photo 2B). Mouth hooks remaining in the mines after each successive molt can be used to identify stages. Larvae lack a typical dorsal plate in the prothorax. Instead they have a dark oblique band that does not cover the dorsal midline.
Pupa
The puparium is oval, slightly flattened ventrally (Photo 2C). Newly formed pupae are greenish and turn dark brown as they mature. Male pupae are smaller (length 4.27 mm and width 1.23 mm) than female pupae (4.67 mm and 1.37 mm).
Adult
Adults are about 1 cm long, with a wingspan of about 1 cm (Photo 2D). There is no obvious sexual dimorphism, although the abdomen in male moths is narrower and pointed posteriorly, while that of females is wider and bulkier. Abdominal scales are gray in males and cream colored in females. Adult moths are mottled gray in color with long and filiform antennae. Taxonomic identification requires dissection of the male genitalia. Care should be taken not to confuse T. absoluta with either the potato tuber moth (Phthorimaea operculella Zeller). Foliar damage by T. absoluta should not be confused with that of Agromyzidae leaf-miner flies such as Liriomyza spp. whose mines in leaves are narrow or small.

Photos 2. The developmental stages of the tomato leafminer, Tuta absoluta: (A) egg, (B) larva, (C) pupa and (D) adult. Photos: Courtesy of CIP.
Biology
Females usually lay eggs on the underside of leaves or stems and to a lesser extent on fruits. Eggs are laid singly (rarely in batches) on all above-ground parts of the host plant. After hatching, larvae enter the plant tissue and begin feeding, thus creating mines. In tomato, young larvae can mine leaves, stems, shoots, flowers, and developing fruit; later instars can attack mature fruit. Larval mines increase in length and width as the larva develops and feeds. Fully-grown larvae usually drop to the ground on a silk thread and pupate in the soil, although pupation may also occur on leaves or in the calyx. Pupation may take place on the surface of the leaf, within the mine or tunnel, or the final instar larva may exit the plant and pupate in the soil, where detection is very difficult. Adult moths are nocturnal; spending the day resting between leaves, and are unlikely to be seen. Adults usually remain hidden during the day, showing greater morning-crepuscular activity. They disperse among crops by flying.
Temperature-dependent development
- absoluta completed its development from egg to adult at temperatures of 10°–32°C (see Annex 7.3.9). Temperatures at 8° and 35°C were lethal for larvae. Tomato leafminer total development was almost eight times longer at 10ºC (164.7 days) than at 32°C (19.4 days). Estimated lower threshold for development of immature stages was 7.9°C (egg), 6.3°C (larva), and 8.6°C (pupa). Mortality was lowest at 20ºC for eggs (5.0%), larvae (10.3%), and pupae (7.1%). Oviposition peaked at 20ºC with 185.8 eggs per female. Population parameters simulated at constant temperatures indicate a population growth between 9.8 to 30.5ºC, with a peak between 24-25ºC.
The established functions (see Annex 7.3.9) were used to estimate the life-table parameters of L. huidobrensis and to build an overall stochastic phenology model. Population parameters indicate that the intrinsic rate of natural increase (rm) augments almost linearly with increasing temperature to reach a maximum at 24°C (0.037) and then sharply decreased at 27°C (0.064). The negative r values below 13°C and above 30°C indicate that population size is decreasing due to high mortality and no or very low reproduction. Similarly, the finite rate of increase for leafminer fly peaked at 24°C (λ=1.082) and was smallest when exposed to 12°C (λ=0.991) and 31°C (λ=0.993); λ values of <1 indicate that the population is decreasing. The highest values for both reproductive parameters (gross reproductive rate and net reproductive rate, Ro) were found at 20°–22°C. The shortest mean generation time (T) and doubling time (Dt) were observed at 30°C (23.7 days) and 24°C (8.7 days), respectively. The optimum temperature for overall population growth ranged from 20°C to 24°C.
Deterministic simulation of life-table parameters under prevailing temperature conditions of two contrasting Peruvian environments showed that lowland (La Molina: 12º 05′ S, 76º 57′ W, 250 masl, with a mean annual precipitation of 6.4 mm and mean temperature of 19.7ºC) present more suitable temperature conditions than do the highland (Huancayo: 12º 00′ S, 75º 22′ W, 3,250 masl, with a mean annual precipitation of 646 mm and temperature of 12.2ºC) for T. absoluta population increase. Life-table parameters as intrinsic and finite rate of increase, as well as net reproductive rate were higher in lowland (Rm: 0.091; λ: 1.096, Ro: 46.4) than in highland (Rm: 0.023; λ: 1.023, Ro: 8.1) environments. On average it will take twice as long to complete one generation in highland (90 days) than lowland (46 days) environments. Further, survival immature life stages were higher in low (72%) than in high (24%) altitude. For the lowland (La Molina) and highland (Huancayo) environments, a mean of 8 and 4 generations per year were determined.
Means of movement and dispersal
Little information exists on the natural dispersal of T. absoluta. However, there are indications that these moths can spread several kilometres by flying or drifting with the wind and can survive in fairly harsh conditions. This suggests that the insect spread may easily occur naturally or be greatly facilitated through agricultural trade.
Economic impact
In South America Tuta absoluta is considered the major limiting factor for tomato production. Larvae of Tuta absoluta attack leaves, buds, stems, flowers, calyces, and tomato fruit. Then severely attacked tomato fruits lose their commercial value. It is a key pest of most greenhouse-grown tomatoes in Argentina and the key pest of tomato production in Chile. Without adequate controls, infestations of T. absoluta can result in 90 to 100 percent loss of field-produced tomatoes in Chile. Also, Tuta absoluta is considered one of the most important lepidopterous pests associated with processing tomatoes in Brazil. Damage to terminal buds in greenhouse-grown hybrid tomato plants in Argentina negatively affects plant architecture and results in reduced plant growth and decreased fruit yield.
Tuta absoluta it is a serious pest of field and greenhouse grown tomatoes in the European and Mediterranean regions. The tomato leafminer caused up to 100 percent losses in tomato crops planted during the winter in the Province of Valencia. Tuta absoluta can complete 9 to 10 generations per year in field tomatoes and 12 generations per year in greenhouse tomatoes. Both yield and fruit quality can be significantly reduced by direct feeding by the larvae, and subsequently by secondary pathogens entering the mines and causing fruit rot. Severely attacked tomato fruits lose their commercial value. Tuta absoluta feeds on potato leaves, including those from Bacillus thuringiensis-transgenic plants, but recent and historical references report that larvae also attack potato tubers.
In Africa, tomatoes are seen as a very promising area for horticultural expansion but the sector is currently experiencing significant impacts from T. absoluta which puts the future of this crop in jeopardy. Losses of up to 80% of the total harvest reportedly led to a three-fold increase in tomato prices in Tanzania in 2016.
Geographical distribution
The tomato leaf miner, Tuta absoluta, was first identified from Peru by Meyrick (1917) from individuals collected in Huancayo (3200 m asl), and has been recorded in all South American countries. The pest is predominantly found in areas below 1000 m above sea level. It was first introduced into Europe in Spain in 2006, from where it is believed to have spread to other European countries and its range has expanded considerably to Asian and African countries in recent years (Fig. 1).
| Africa | Algeria, Angola, Botswana, Burkina Faso, Cape Verde, Egypt, Eritrea, Ethiopia, Kenya, Ghana, Kenya, Libya, Malawi, Mayotte, Morocco, Mozambique, Namibia, Niger, Nigeria, Rwanda, Senegal, Spain (Canary Islands), South Africa, Sudan, , Tanzania, Tunisia, Uganda, Zambia, Zimbabwe |
| Asia | Armenia, Azerbaijan, Bangladesh, India, Iran, Iraq,, Israel, Jordan, Kyrgyzstan, Nepal, Qatar, Saudi Arabia, Syria Arab Republic, Turkmenistan, United Arab, Emiratos, Yemen |
| Europe | Albania, Austria (under eradication), Belarus, Belgium, Bosnia and Herzegovina, Bulgaria, Croatia, Cyprus, Czech Republic (under eradication), France, Georgia, Germany (under eradication), Greece, Guernsey, Hungary, Italy, Kosovo, Lithuania, Malta, Montenegro, Netherlands, Portugal, Romania, Russia, Serbia, Slovenia, Spain, Switzerland, Turkey, Ukrania (under eradication) |
| Central America and the Caribbean | Costa Rica, Panama |
| South America | Argentina, Brazil (Goias, Minas Gerais, Sao Paulo), Bolivia, Cayman Islands, Chile, Colombia, Ecuador, Paraguay, , Peru, Uruguay, Venezuela |
| Oceania | Australia (absent, intercepted only) |
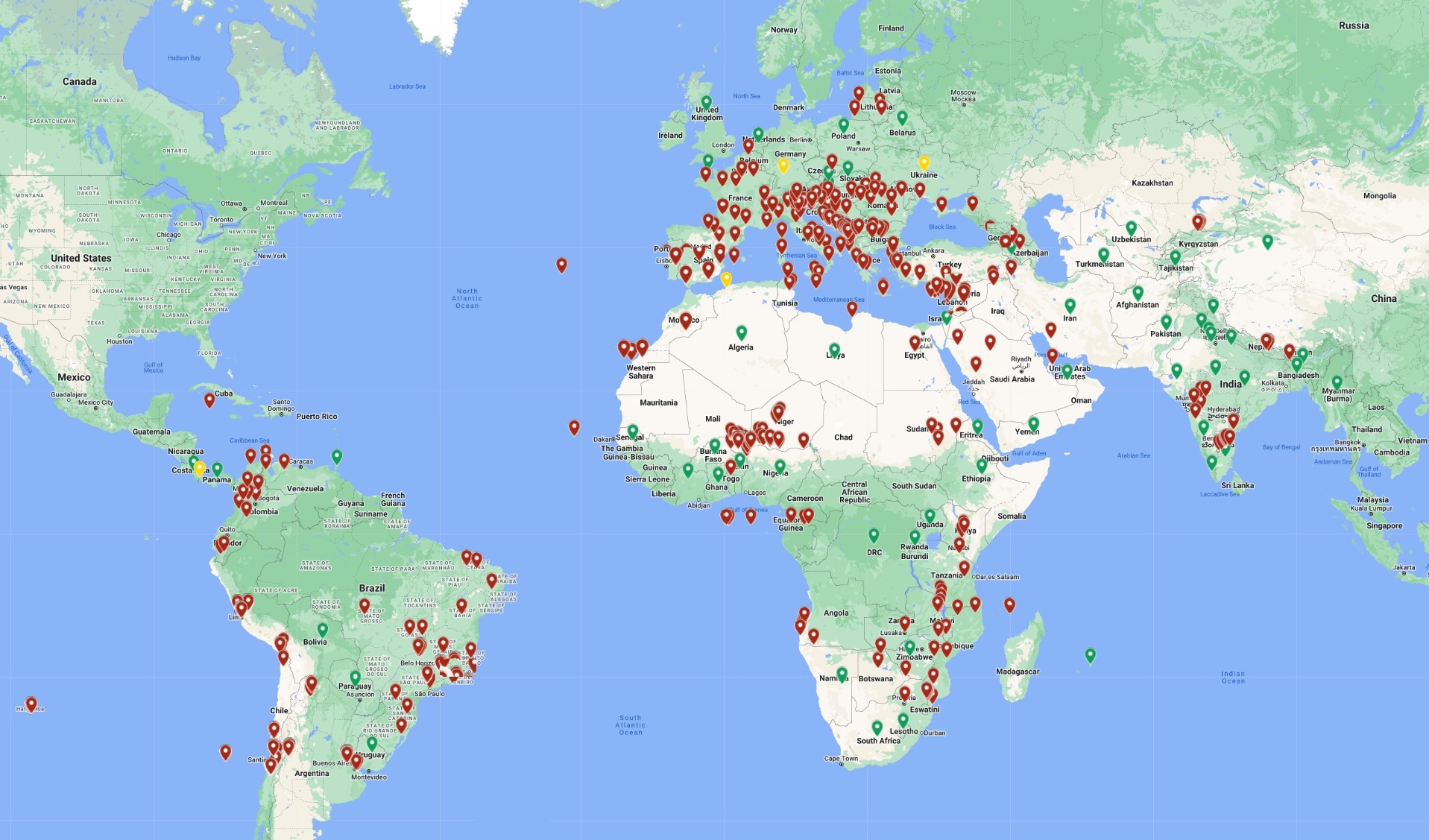
Figure 1. Global geographical distribution of the serpentine tomato leafminer, Tuta absoluta. Green points indicate countries with reported pest establishment; yellow points are countries with reported occurrence in protected crops (greenhouses); and red points are georeferenced distribution data.
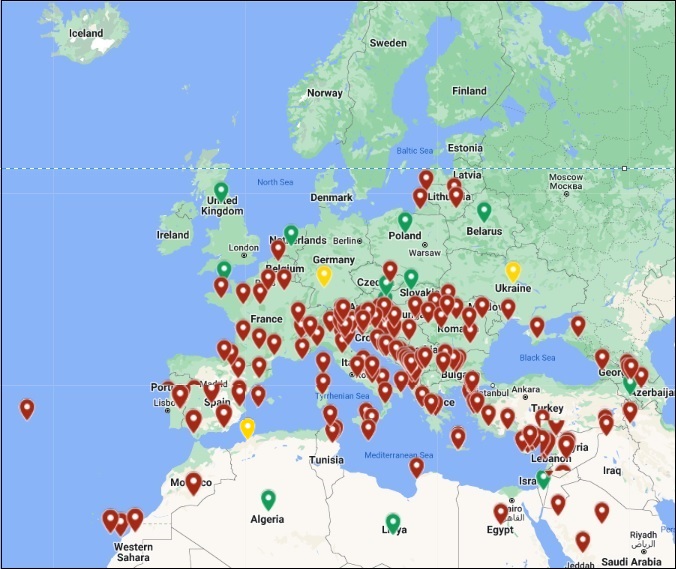
Figure 1. Geographical distribution of the serpetine tomato leafminer, Tuta absoluta, in Europe.
Phytosanitary risks
Tuta absoluta is potentially a serious pest of tomato in warmer parts of the world, both in the field and in protected conditions. In Europe it has become a major pest of tomato grown under glass, and is officially listed as an A1 pest since its introduction into several European countries. A pest risk analysis carried out by the CGIAR Research Program on Roots, Tubers, and Banana (RTB) for Eastern African countries in 2015 defined some pathways in which T. absoluta can entry and spread in some African countries: Fruits or tubers imported from countries where the pest is present (tomato: very likely with high confidence; potato: very unlikely with medium confidence), plants for planting, packing material and transportation vehicle, cross border trade of tomato fruit/ potato tubers and passenger luggage. In China, the following potential invasion pathways were defined: (a) Long-distance transmission: international agricultural trade, tomato fruits and plants, production facilities and packages, and transportation vehicles; (b) Short-distance dispersal: Natural factors (wind/water) and Larva crawling and adult flight. Most likely pathways for introduction of T. asboluta are international traffic (fresh tomatoes), border trade points (tomato fruits and plants) and unofficial introduction (tomato fruits and plants)
Risks mapping under current and future climates
Global Risks
Changes in establishment and future distribution
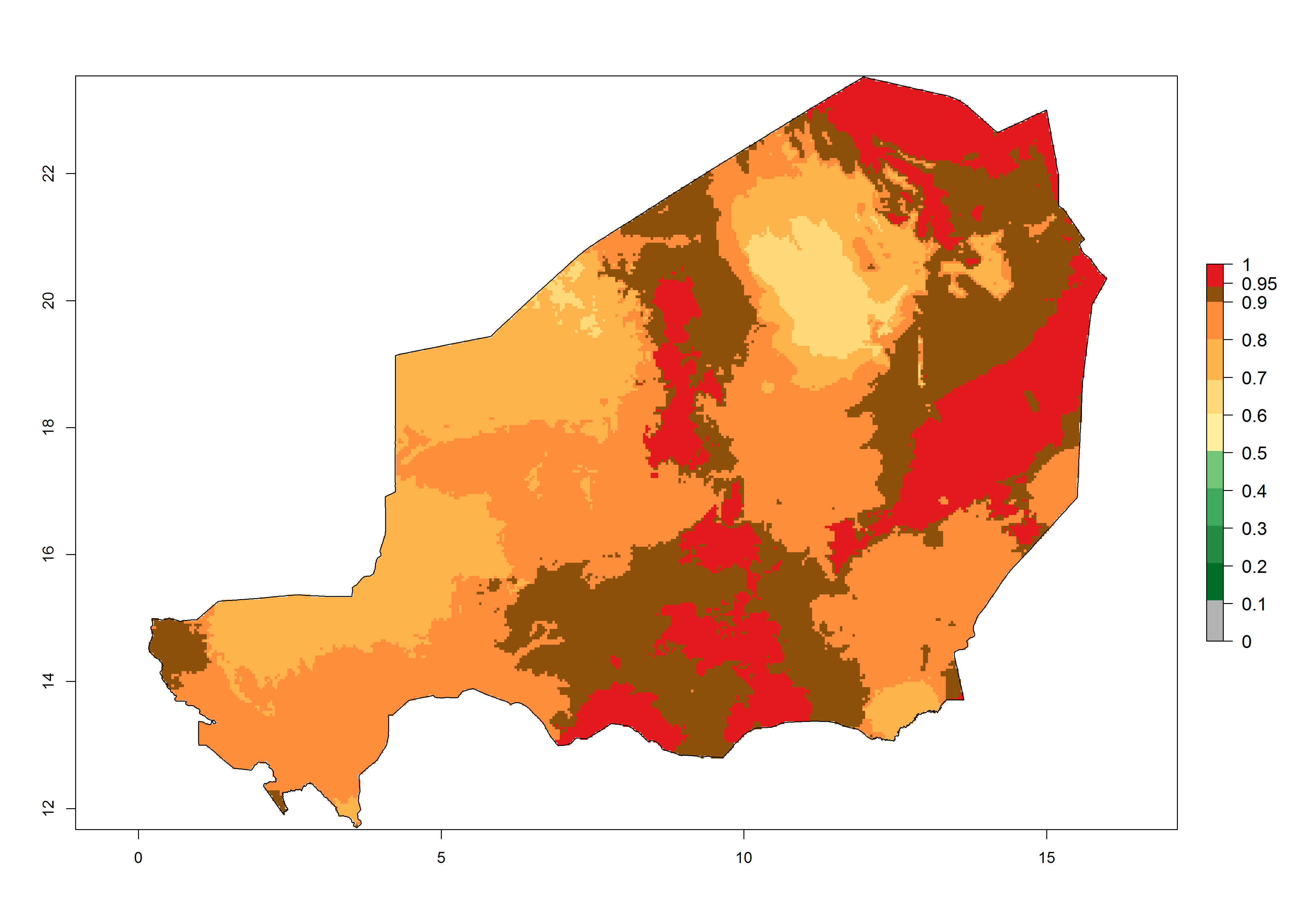
An establishment risk index (ERI)>0.95 reflects well the current global distribution of T. asboluta under the current climate of the year 2000 (Fig. 2). Those regions include Central and South America; Africa (Northern, Eastern, western, central and southern countries); Southern Asia countries as Bangladesh, Nepal and southern India (Karnataka, Maharashtra and TamilNadu); Middle East (Arabia Saudi, Yemen); and southern European countries such as Portugal, Spain, Italy, Turkey (compare with Fig. 1).
A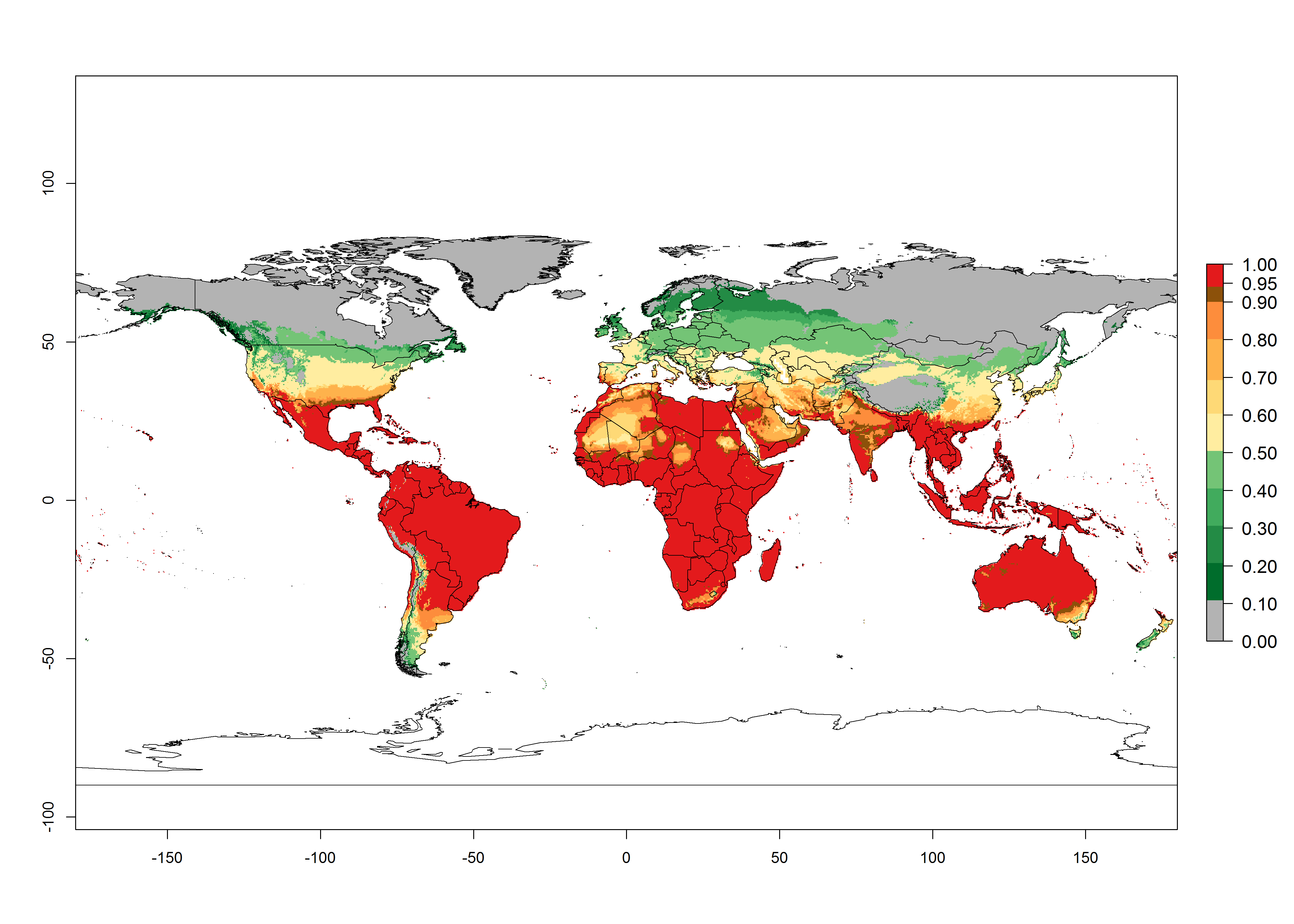 |
B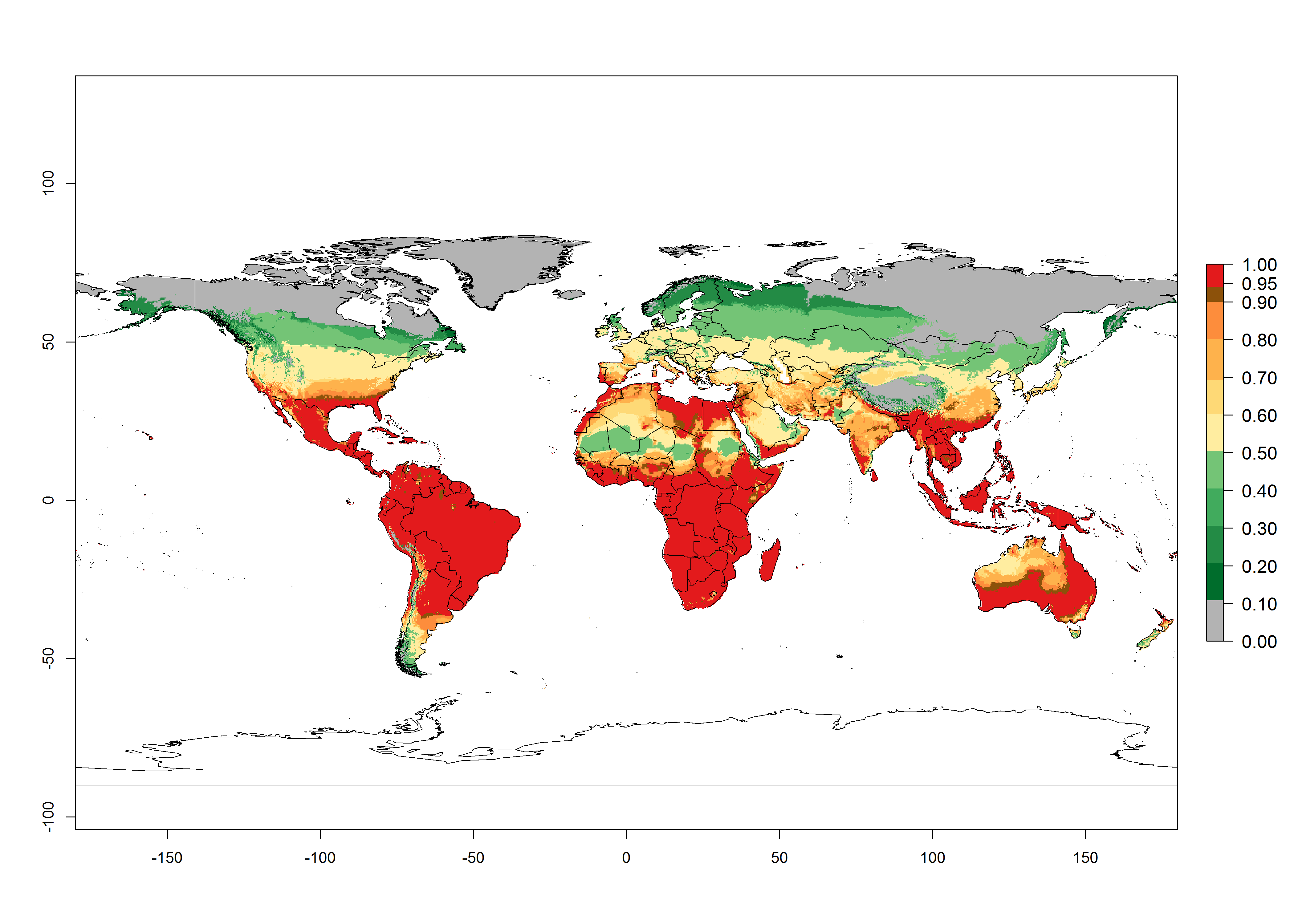 |
C 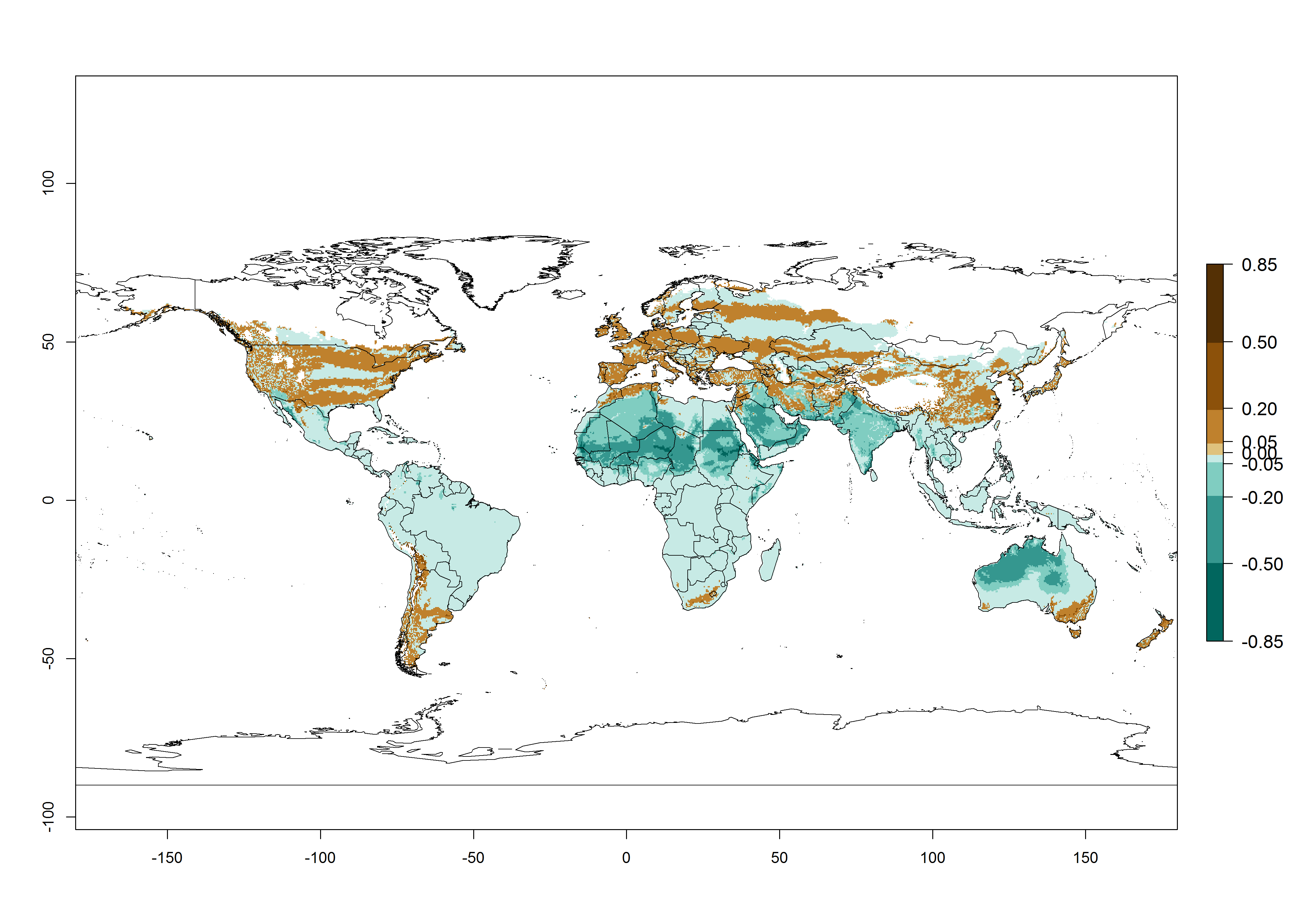 |
Figure 2. Changes in establishment and potential distribution of the tomato leafminer, Tuta absoluta, worldwide according to model predictions, using the ERI for the years 2000 (A) and 2050 (B), and changes of the ERI between 2000 and 2050 (C). An ERI>0.7 is associated with potential permanent establishment.
In zones where the ERI drops below the maximum number of 1, the likelihood of long-term establishment is reduced, but T. asboluta also occurs in regions with an ERI of 0.7–0.9 (light- and dark-orange zones) as in Middle East (Syria, Jordan, Iraq), Southern Asia (Iran), and some areas of southern European countries (Greece, France, Portugal, Spain, Albania, Italy). T. asboluta has also been reported in regions with a low likelihood of establishment (ERI<0.6) as in temperate regions of Northern (Lithuania), Eastern (Belarus, Bulgaria, Czech Republic, Hungary, Romania) and Western (Austria, Belgium, The Netherland) European countries; and some countries of Middle East (Georgia, Armenia, Azerbaijan). Under these conditions, T. asboluta survives unfavorable conditions under natural conditions or continues its development in protected production systems. Reported occurrences are due to survival and infestations in greenhouses, not to survival under open-field conditions.
Global predictions for 2050 indicate that T. asboluta will continue to be a high-risk pest in most of the tropical and subtropical areas (Fig. 2B). However, a decrease in the high risk of establishment (ERI>0.8) is predicted in South Asia (India); regions in North and West Africa; Middle East (Saudi Arabia and Oman); and Australia. A slight range expansion (+0.20) is observed in more temperate areas such as western, central, and eastern Europe, North America (northern U.S. and southern Canada), and western Pacific (Japan and Korea) (Fig. 2C), but still with a low establishment potential of the pest (ERI<0.6). Further, it is predicted that T. asboluta will potentially shift further into southern South America (Bolivia, Chile and Argentina).
Changes in abundance
Figures 3A and B illustrate the mean numbers of generations that might theoretically develop worldwide within a year in the years 2000 and 2050 scenarios. Global maps of the generation index (AI) in year 2000 estimates between 11-17 generations per year in most subtropical and tropical regions. For temperate zones is predicted between 3-8 generations per year. The generation index (GI) change indicates a potential increase of 2–3 generations per year in some areas de Central (Mexico, Guatemala, Honduras, El Salvador, Nicaragua, Costa Rica, Panama) and South (Venezuela, Colombia, Ecuador, eastern coast of Brazil, northern Chile, coast of Peru) America, central (Angola, Congo, Gabon, Cameroon) and eastern (Ethiopia, Rwanda, Burundi, Tanzania, Zambia, Malawi, Zimbabwe) and coast of northern (Morocco) African countries, and Middle East (Yemen). In temperate regions only a slight increase of 1–2 generations per year is expected. By contrast, a decrease of 1 generation per year is estimated for some areas in the Middle East (Arabia Saudi) and African countries (Mauritania, Malia, Niger, Chad and Sudan).
Globally, simulated GIs gave reasonable predictions when compared with generation numbers reported in the literature for South America. For example, in lowland regions as the Cañete valley (Peru), Apaza valley (Chile) and Pernambuco (Brazil), 9 (GI=8-11), 7-8 (GI=5-8), and 9 (GI=8-11) generations per year, respectively, have been reported. In the temperate zone of Buenos Aires (Argentina) were registered 5 (GI=5-8) generations per year. Estimations made by the GI are consistent with these data.
Peruvian localities of La Molina (central lowland, 500 masl, mean annual temperature of 20ºC) and Huancayo (central highland 3,300 masl, mean annual temperature of 12ºC), mean numbers of 12 and 7 generations per year, respectively, have been reported. Estimations made by the GI are consistent with these data. Change of T. absoluta GI between 2000 and 2050 scenarios shows that the number of tomato leafminer generations per year will increase more rapidly in lowland (2–3 more generations per year) in comparison with temperate zones with a predicted increase of less than 1-2 generation per year.
Global maps of the activity index (AI) in year 2000 estimates a high activity of T. absoluta by a factor of 13–21 in most tropical regions, and by a factor of 7–13 for most subtropical regions of South America, East and Central Africa, and Southeast Asia (Fig. 3D). Lowest activity is guesses for temperate zones (AI=1-7). Predictions of potential changes for the 2050 temperature scenario indicate a strong decrease in the potential growth of T. absoluta in tropical regions of Central and South America, Africa, and Asia and a moderate decrease in subtropical areas (Fig. 3D, F). In most of the temperate regions and subtropical areas of South America and Africa, the AI increases from the years 2000–2050 by values up to 1.
| GI | AI | |||
| 2000 |
|
|
||
| 2050 |
|
|
||
| Index change (2000 – 2050) |
|
|
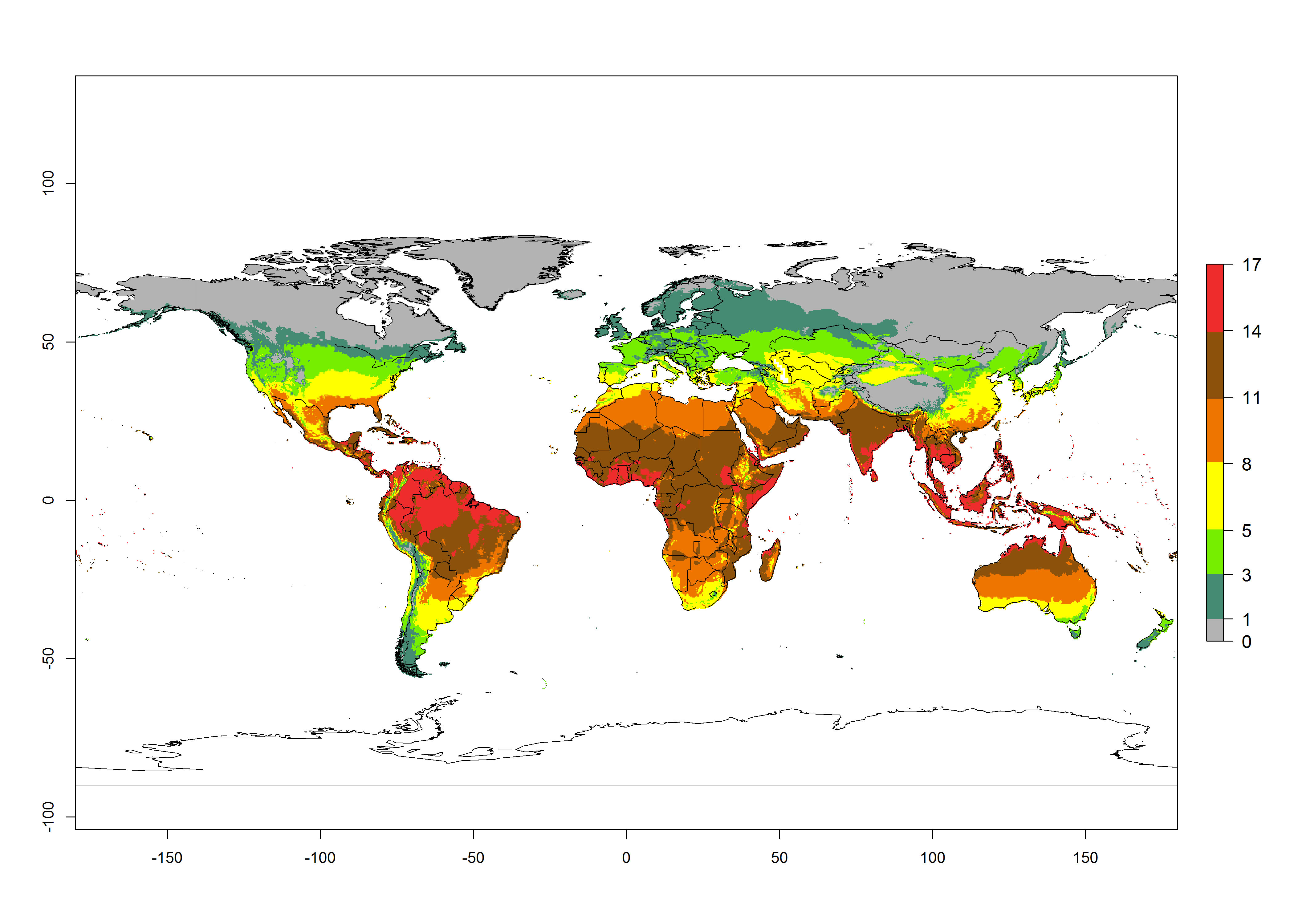
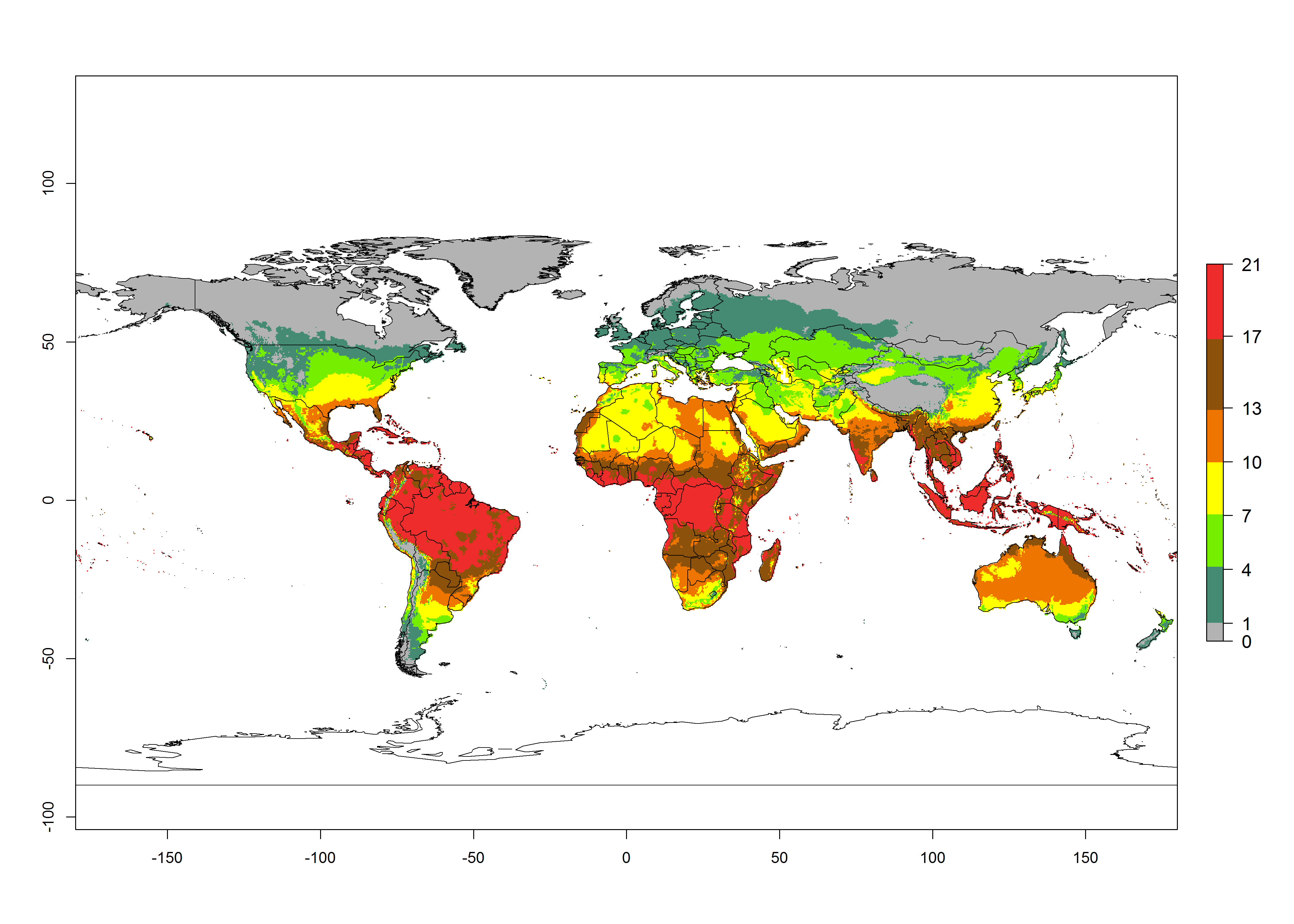
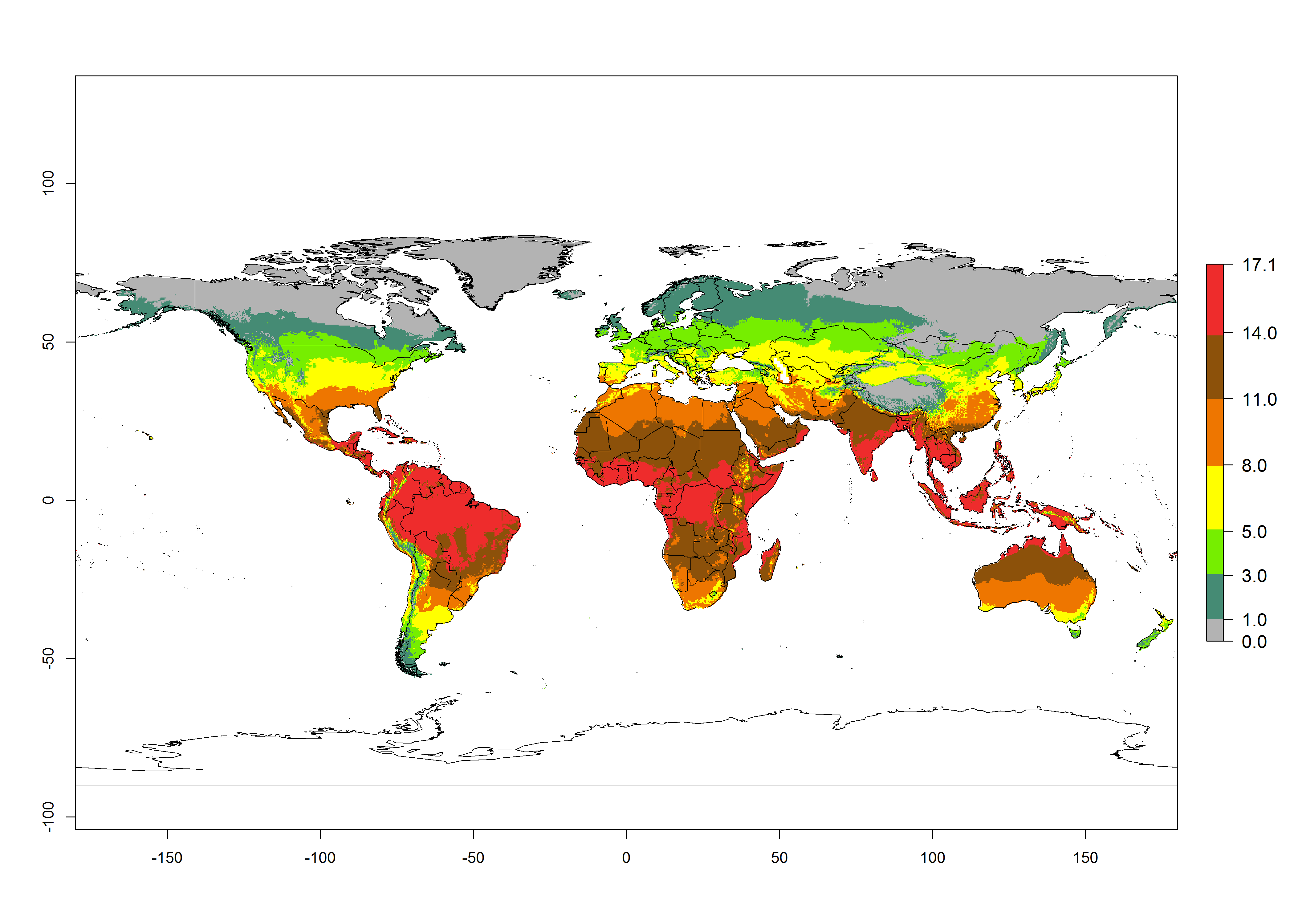
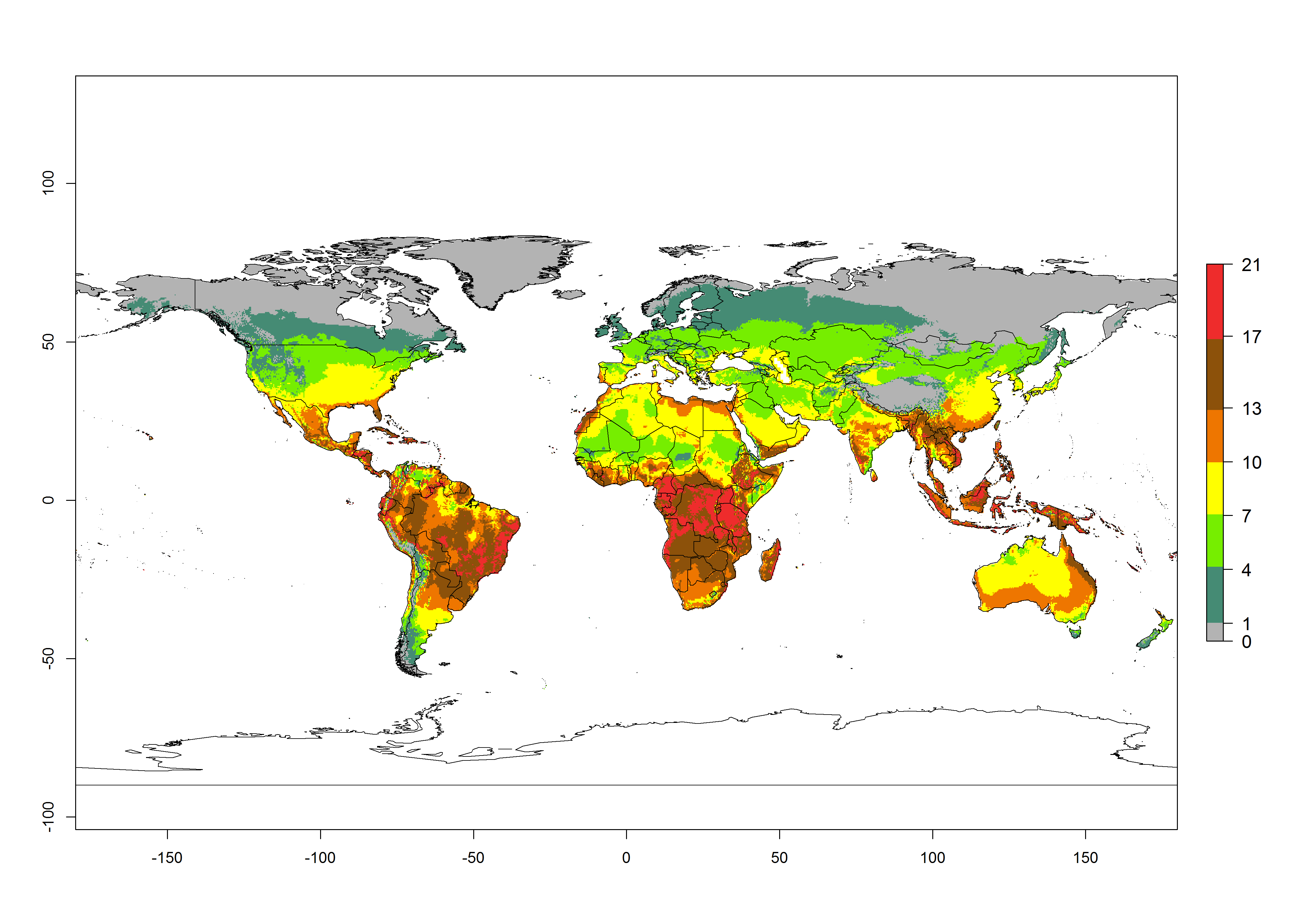
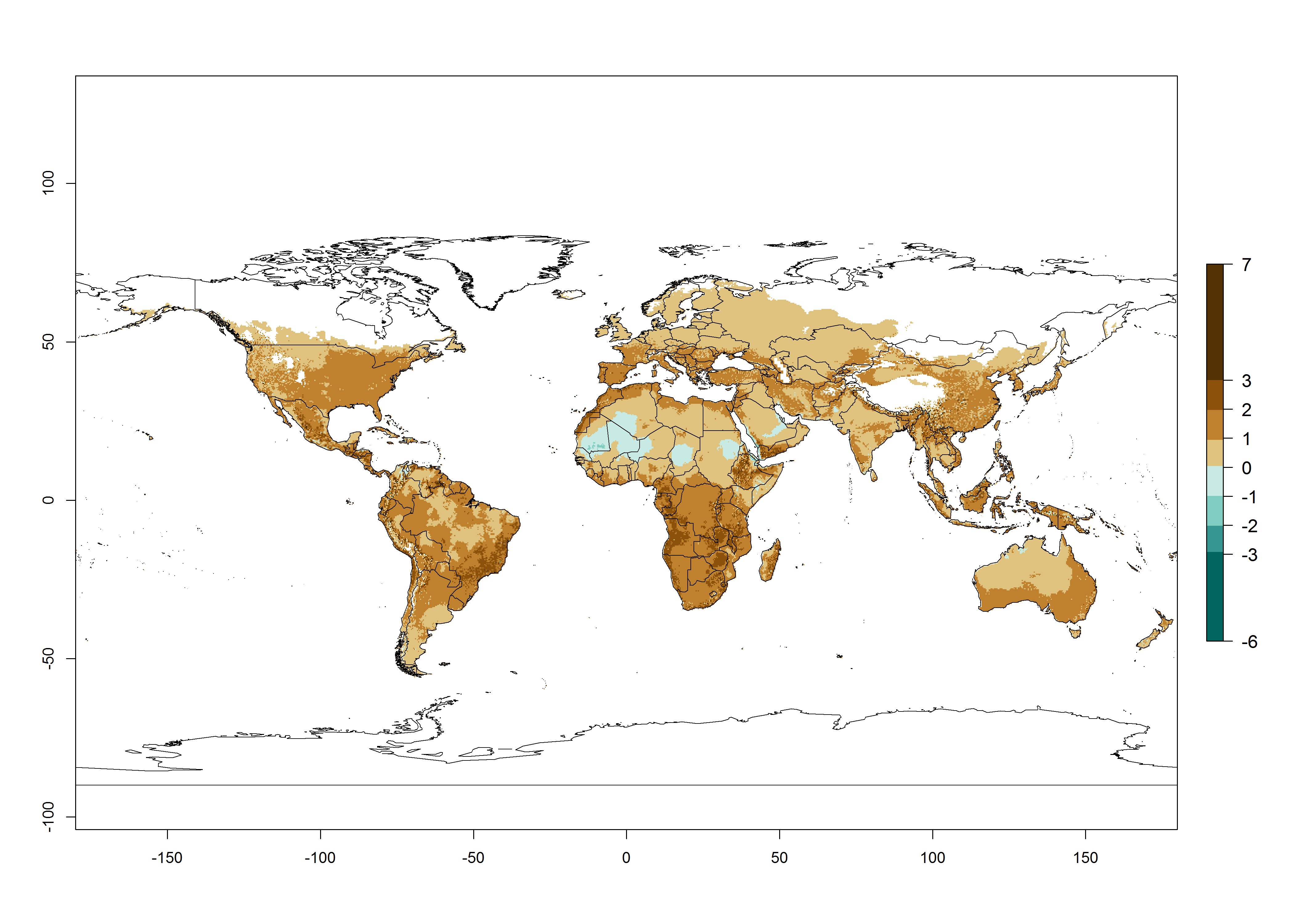
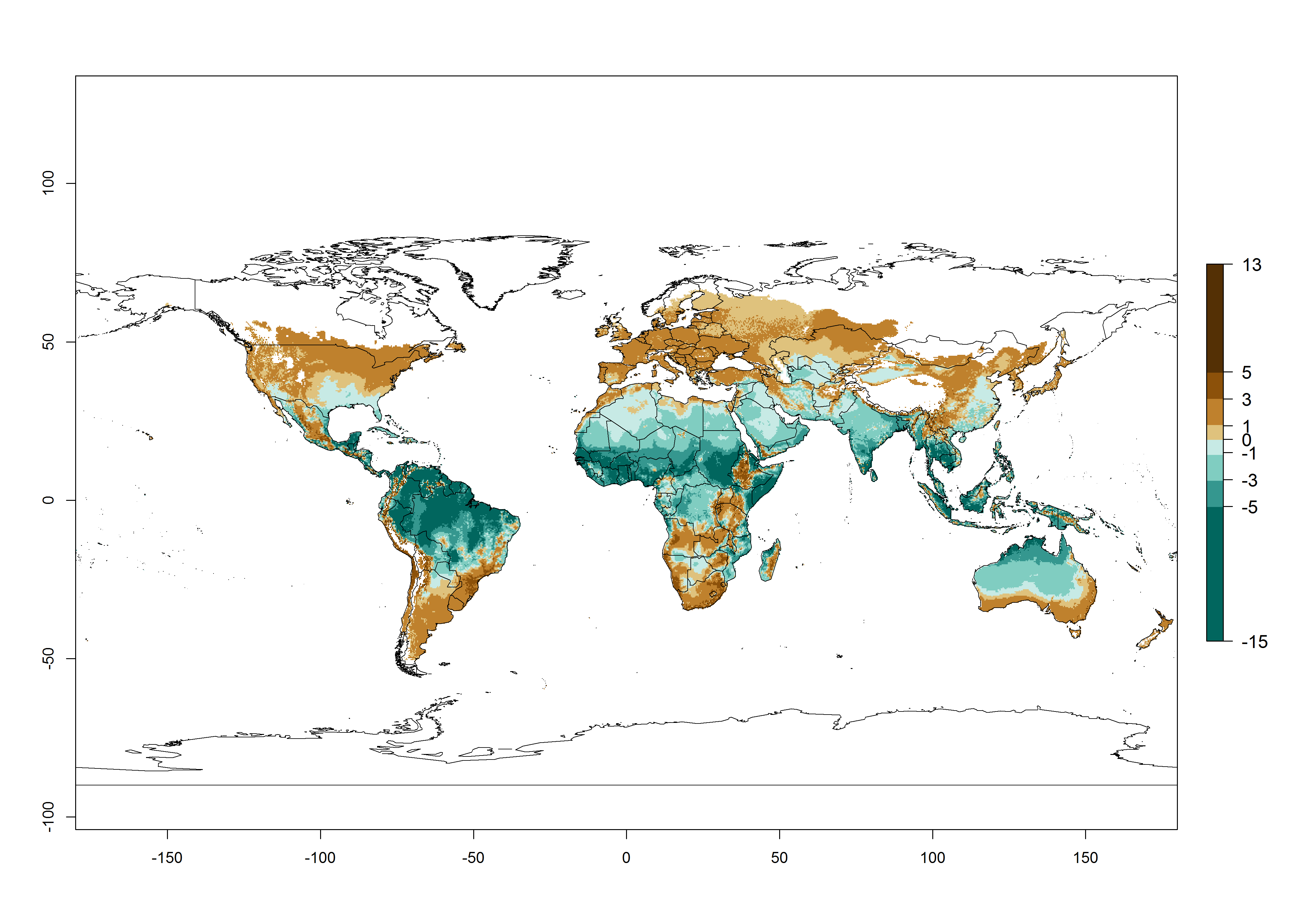
Figure 3. Changes in abundance (generation index [GI], damage potential) and activity (activity index [AI], potential population growth) of the tomato leafminer, Tuta absoluta, worldwide according to model predictions, using the GI (A, B) and the AI (D, E) for the years 2000 and 2050, and the absolute index change (C, F).
Regional Risks for Africa
Changes in establishment and future distribution
In Africa, T. absoluta is already present in most Eastern, Central, Northern, Western and Southern African countries with an ERI>0.95 (Fig. 4A). However, T. absoluta is already distributed under open-field conditions in regions with an ERI>0.7–0.9 (light- and dark-orange zones) as in some areas of Niger and South Africa. Under the year 2050 temperature scenario, a slight decrease (<-0.05) in most of the eastern, central, and southern regions is observed, but T. absoluta will remain a potential risk and threat for food production. A high decrease (-0.2 to -0.5) in the risk of establishment is predicted in most of the countries associated with Sahara desert, mainly in Mauritania, Mali, central Chad and northern Sudan where high temperatures will be unfavorable for the development of T. absoluta (ERI<0.5). By contrast, a moderate increase in the spread of T. absoluta (0.0.5–0.2) with a high establishment potential (ERI>0.7) is predicted for some areas of South Africa and Mediterranean countries (Morocco and northern Algeria) (Fig. 4B, C).
A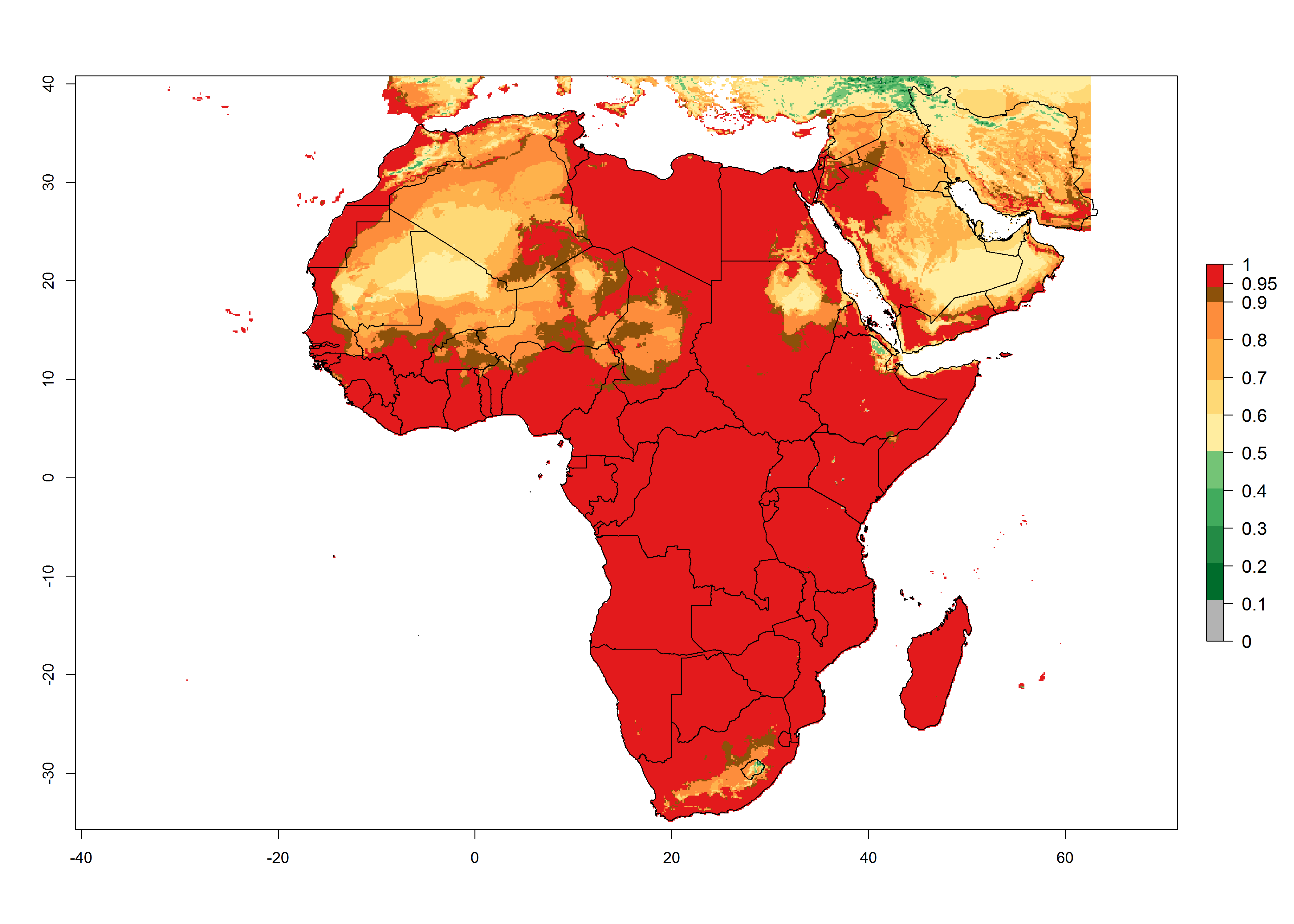 |
B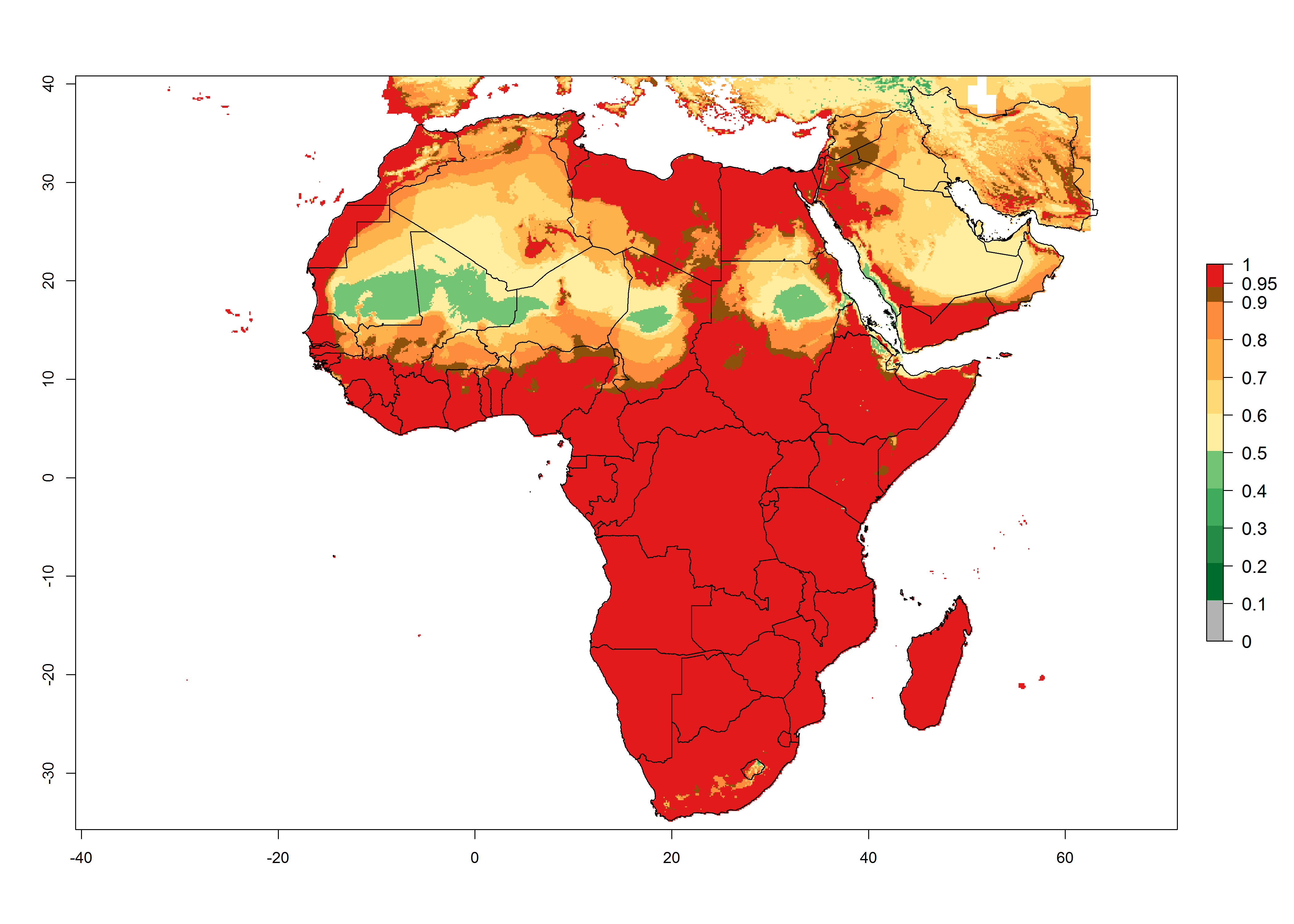 |
C 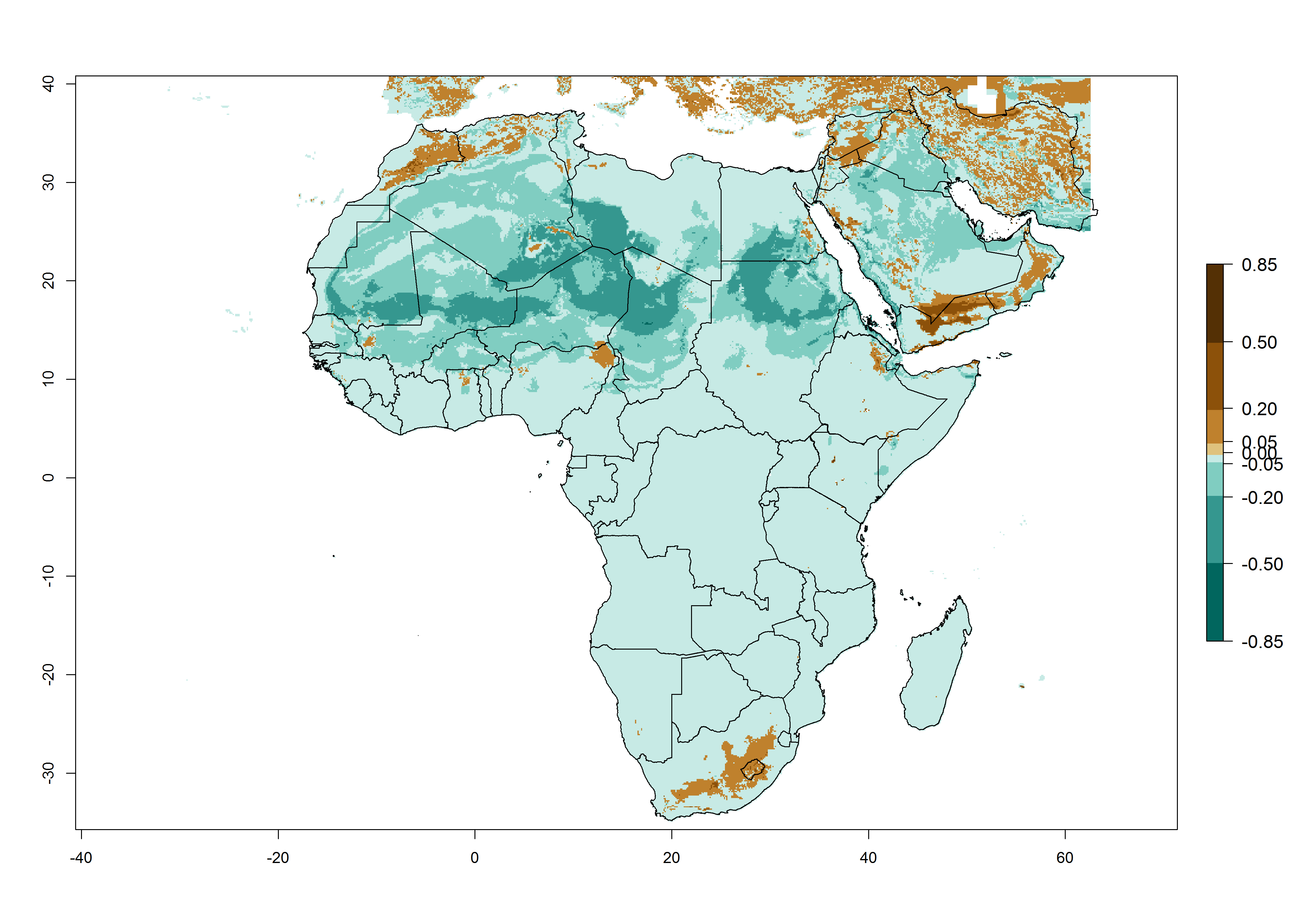 |
Figure 4. Changes in establishment and potential distribution of the tomato leafminer, Tuta absoluta, in Africa according to model predictions, using the ERI for the years 2000 (A) and 2050 (B), and changes of the ERI between 2000 and 2050 (C). An ERI>0.7 is associated with potential permanent establishment.
Changes in abundance
Under the present climate scenario, the highest GI indicates 14–17 generations of T. absoluta per year in some of Eastern (Somalia, Ethiopia, Kenya, and Sudan) and western (mainly Nigeria, Benin, Togo, Ghana, and Cote d’Ivoire) African countries (Fig. 5A). In central Africa is expected between 11-14 generations per year. The lowest GI (5-8 generations per year) is estimated for Morocco, northern Algeria and South Africa. Under the year 2050 temperature scenario the maximum abundance (14–17 generations per year) of T. absoluta is estimated in Western, Central and Eastern Africa (Fig. 5C). The map of absolute index change shows an increase in the number of T. absoluta generations (1–2 per year) for most central, eastern and southern African countries (Fig. 5C). In the case of activity index (AI), the highest activity by a factor of 17-21 of T. absoluta is concentrated in the western, eastern and central regions of Africa (Fig. 5D). An increase in the temperature under the year 2050 scenario predicts a decrease (-1 to -5) in these regions but T. absoluta will remain a high population growth potential (AI>17) (Fig. 5E). It is estimated that in Africa the change of activity between 2000 and 2050 will be increase by a factor of 1–3 in Angola, Zambia, Ethiopia, Namibia, Botswana, Zimbabwe, South Africa, and the Mediterranean coast of Morocco and Egypt.
| GI | AI | |||
| 2000 |
|
|
||
| 2050 |
|
|
||
| Index change (2000 – 2050) |
|
|
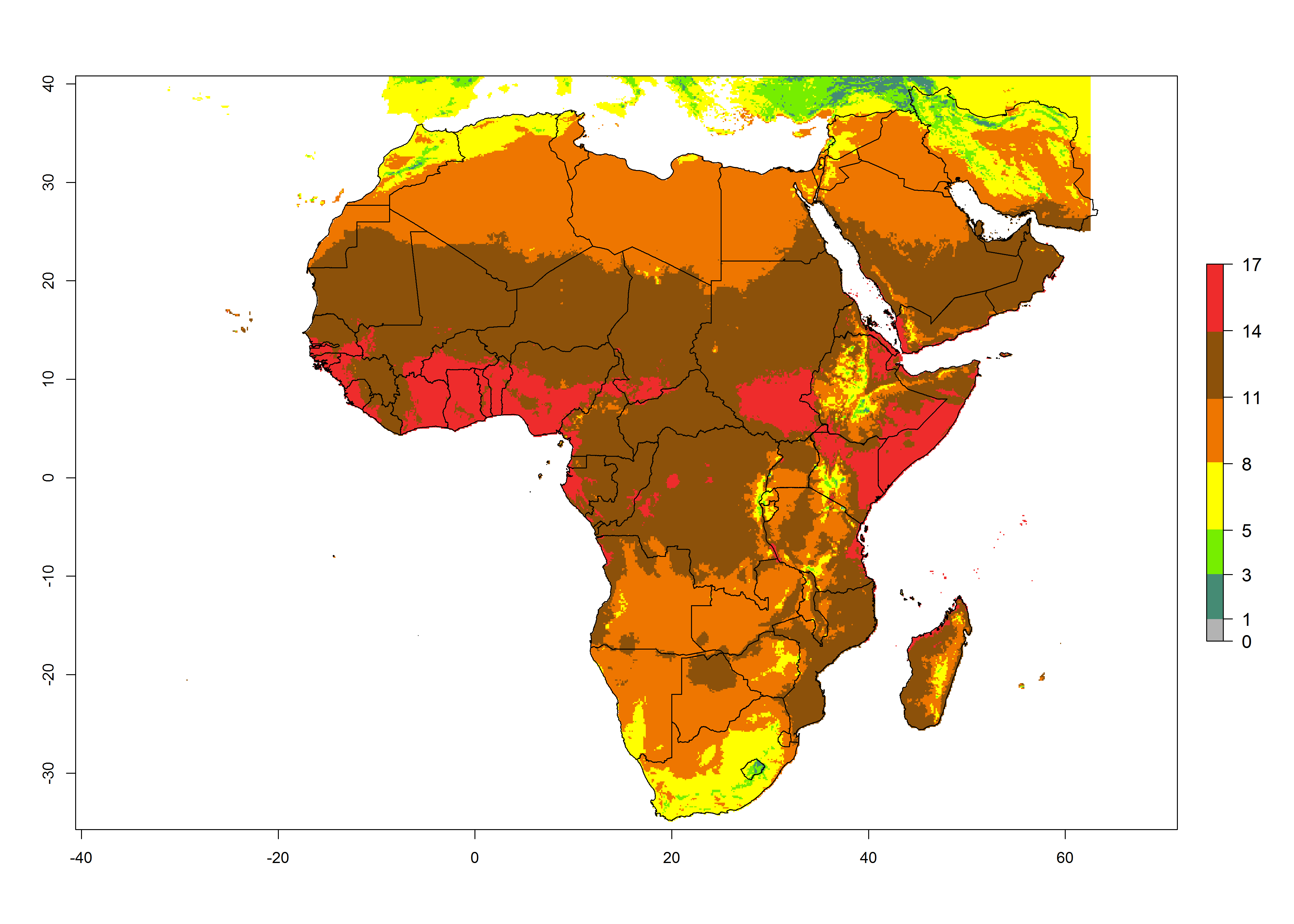
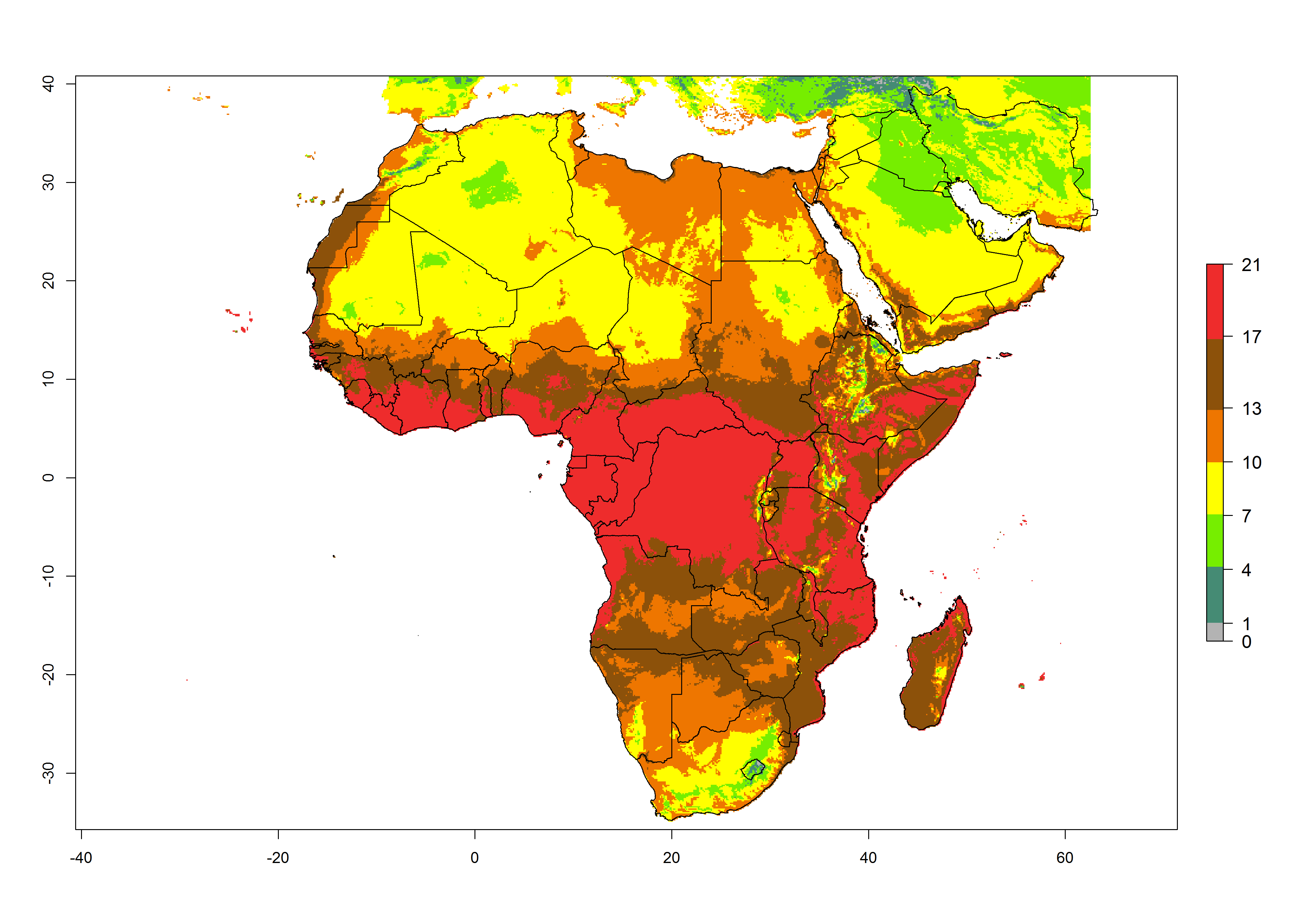
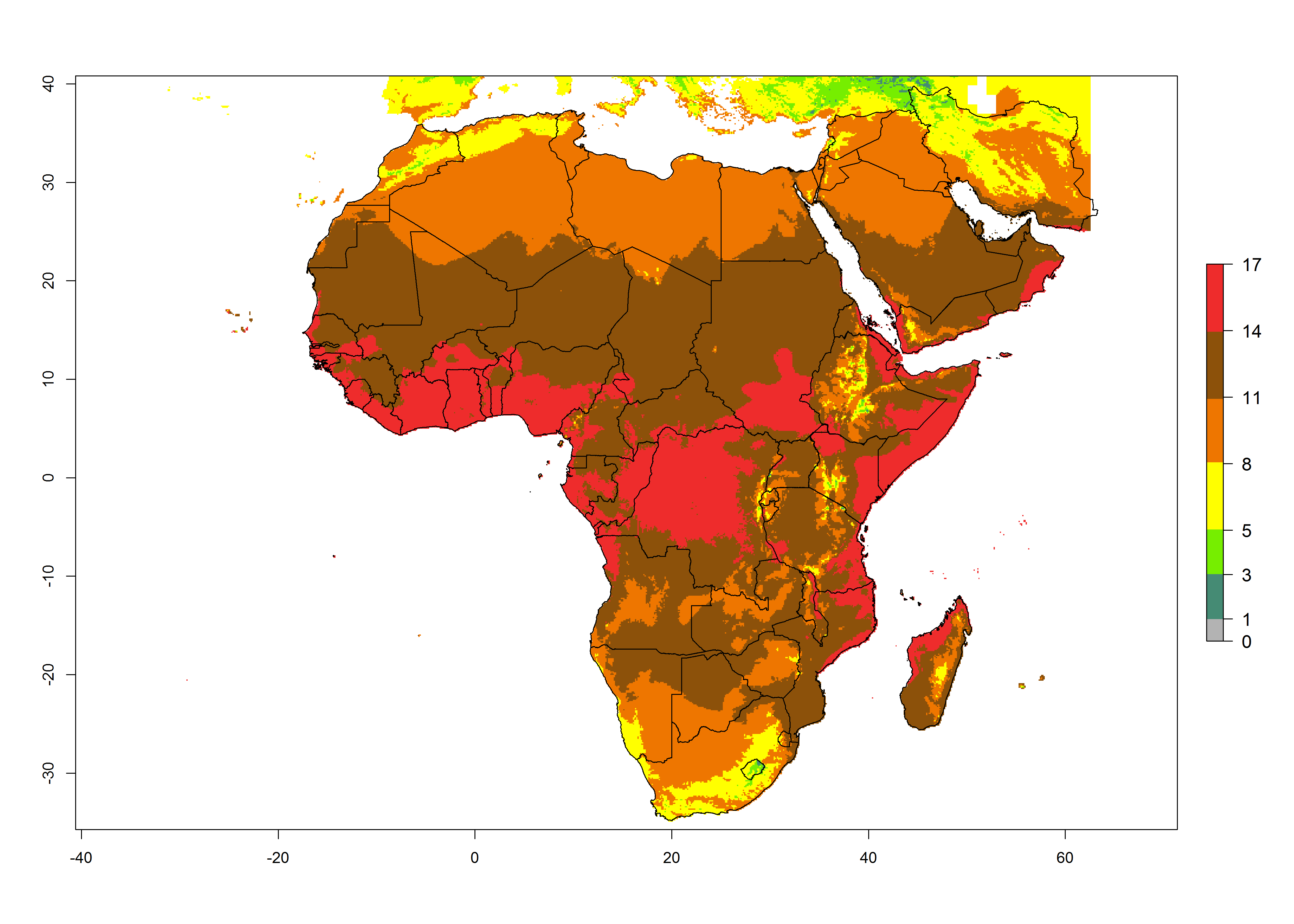
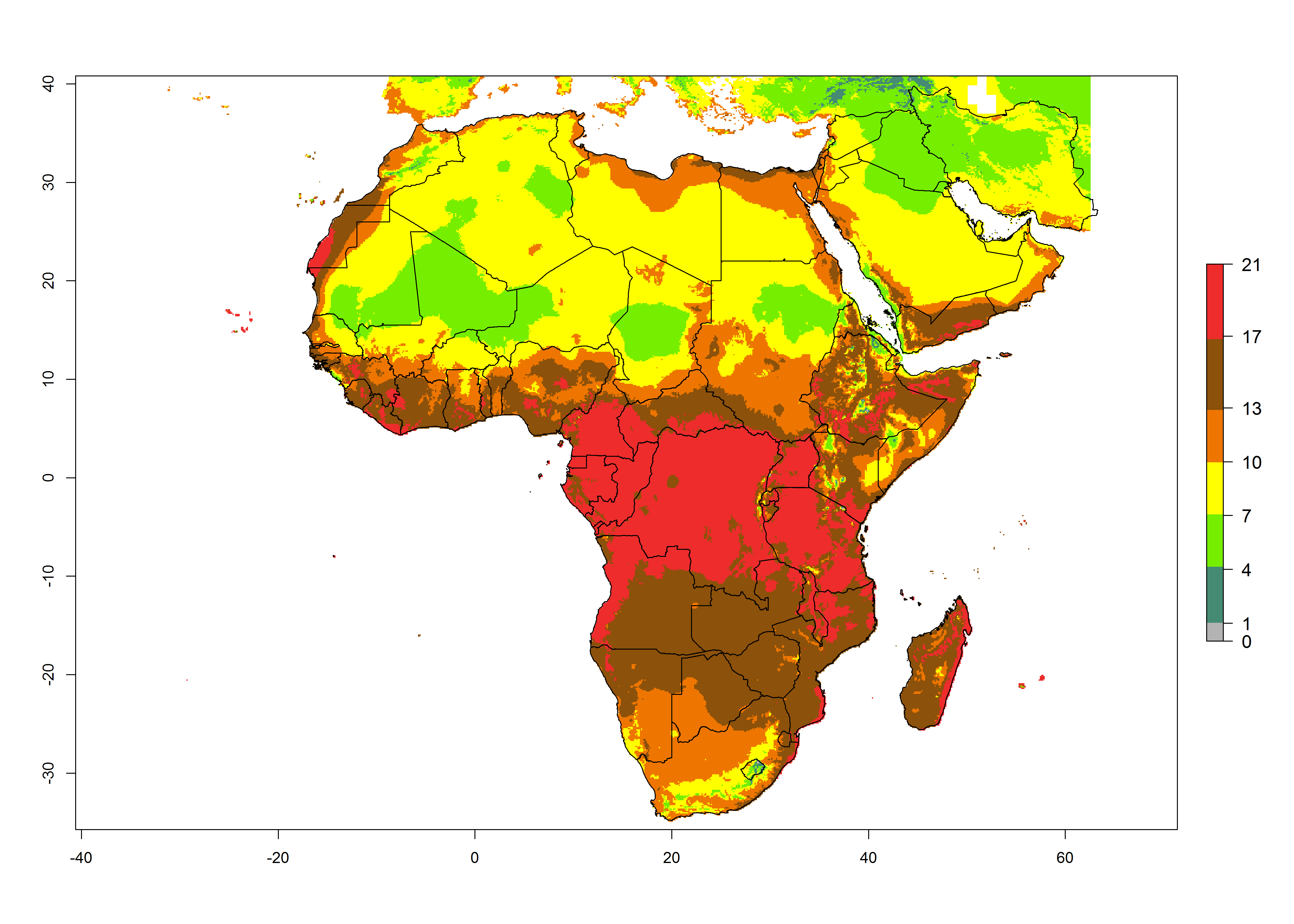
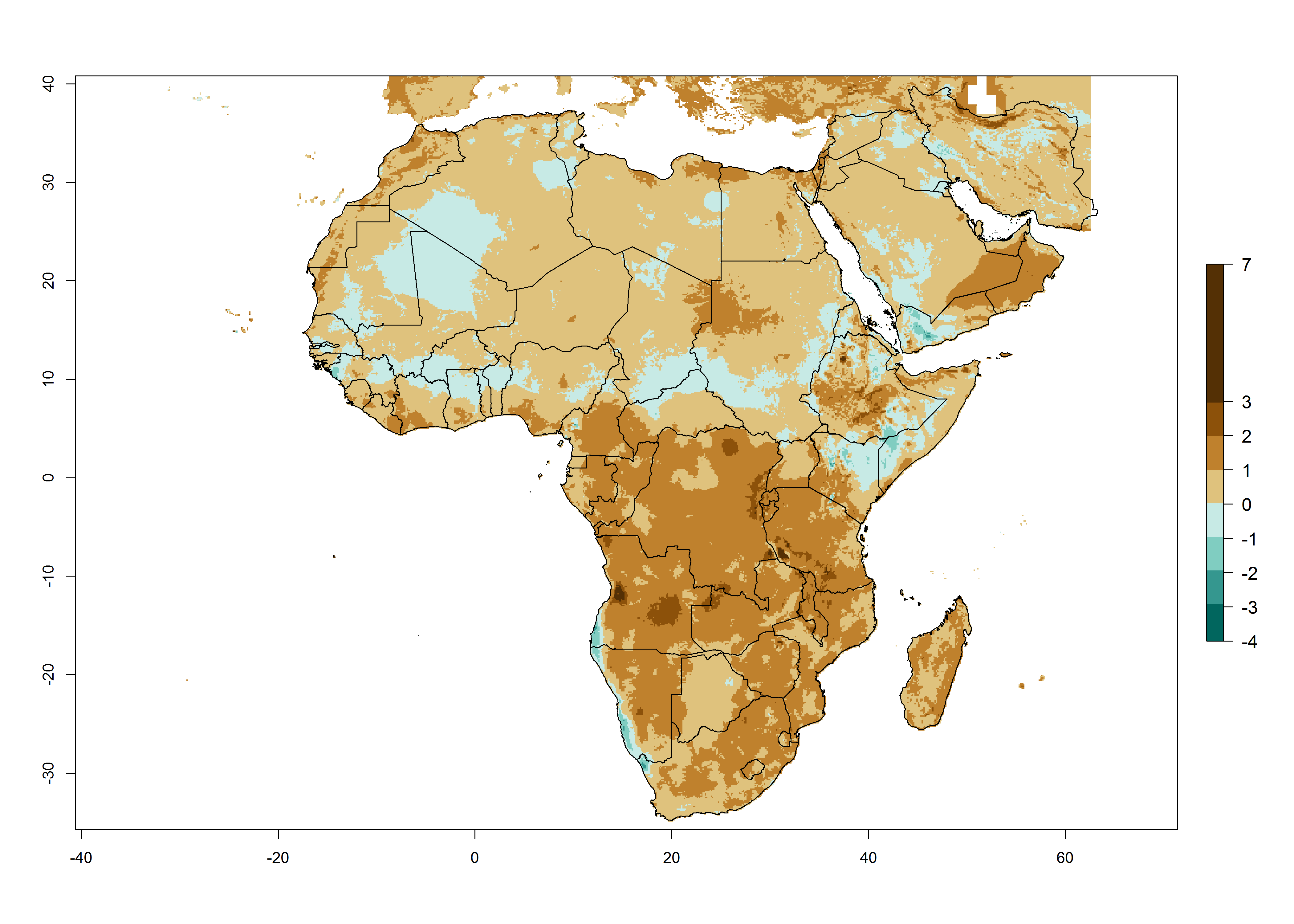
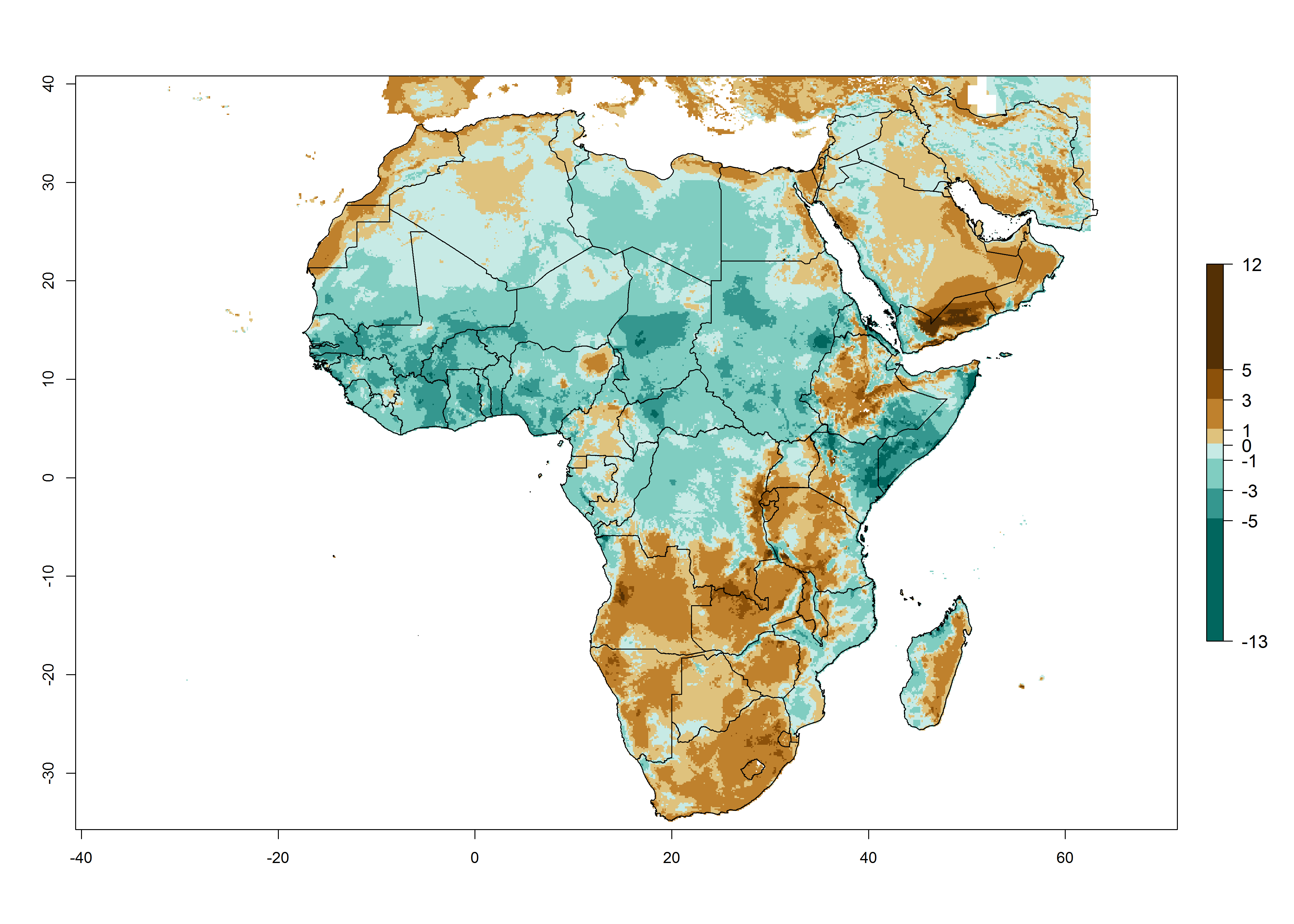
Figure 5. Changes in abundance (GI, damage potential) and activity (AI, potential population growth) of the tomato leafminer, Tuta absoluta, in Africa according to model predictions, using the GI (A, B) and the AI (D, E) for the years 2000 and 2050, and the absolute index change (C, F).
Country Risk Maps
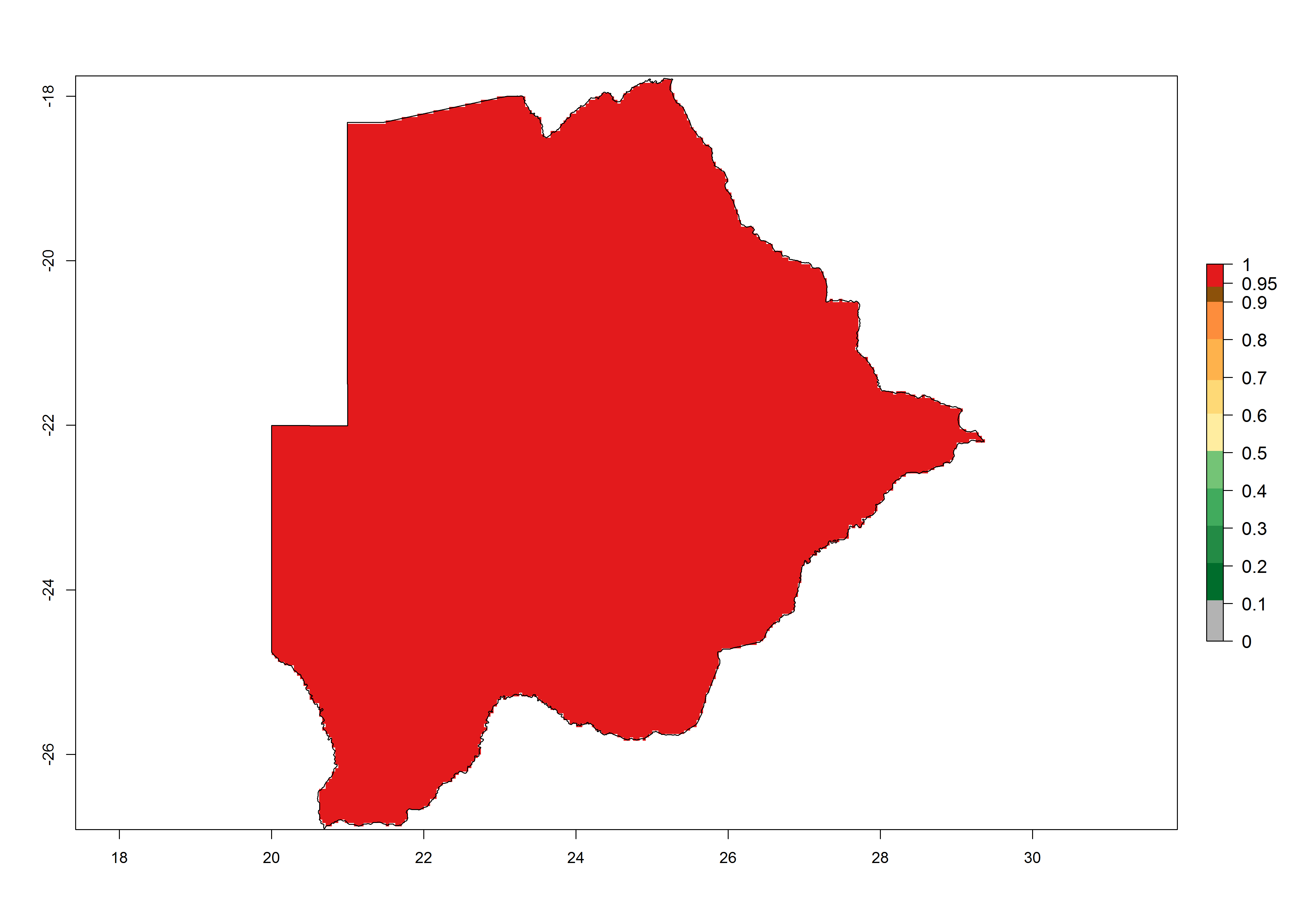
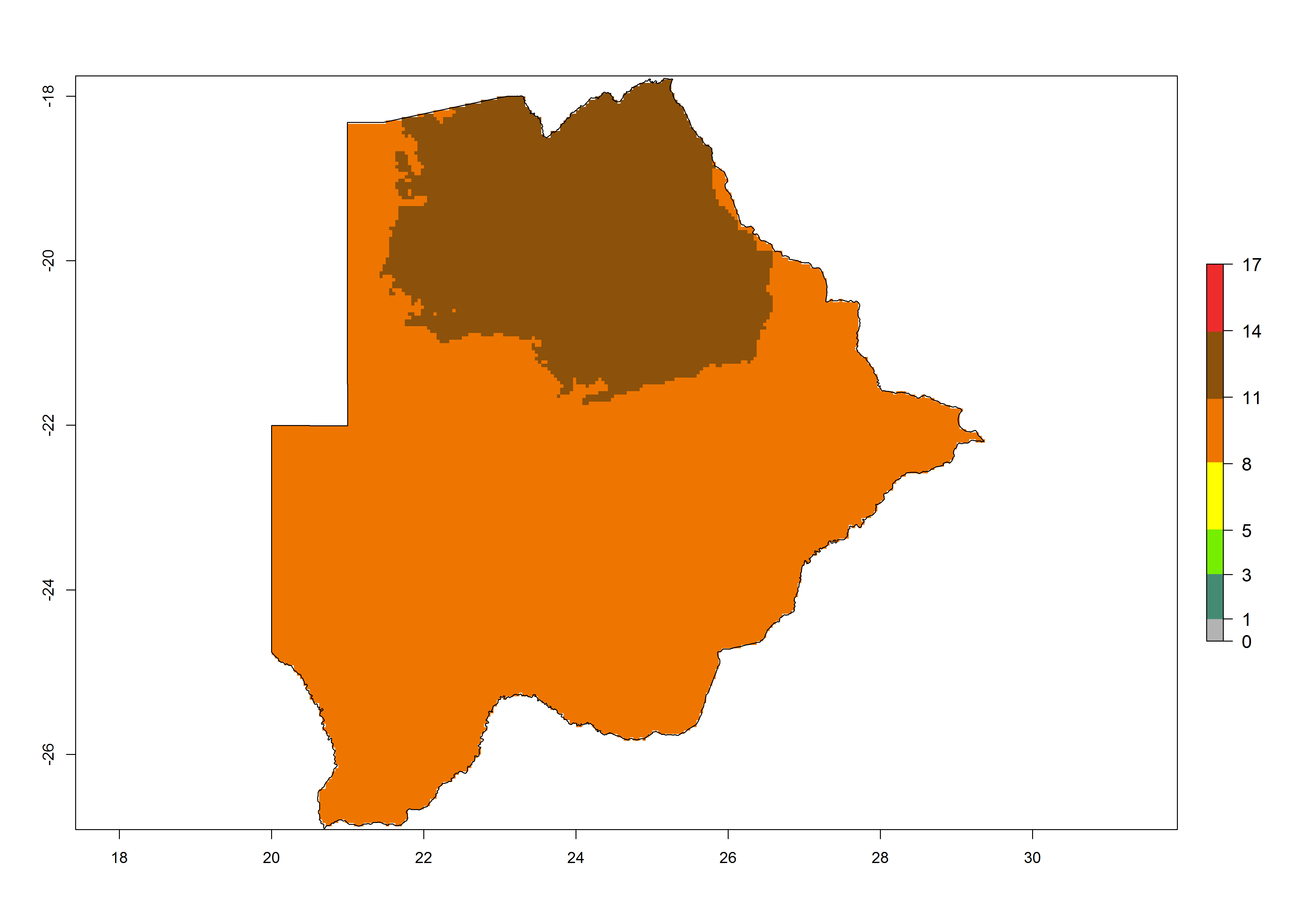
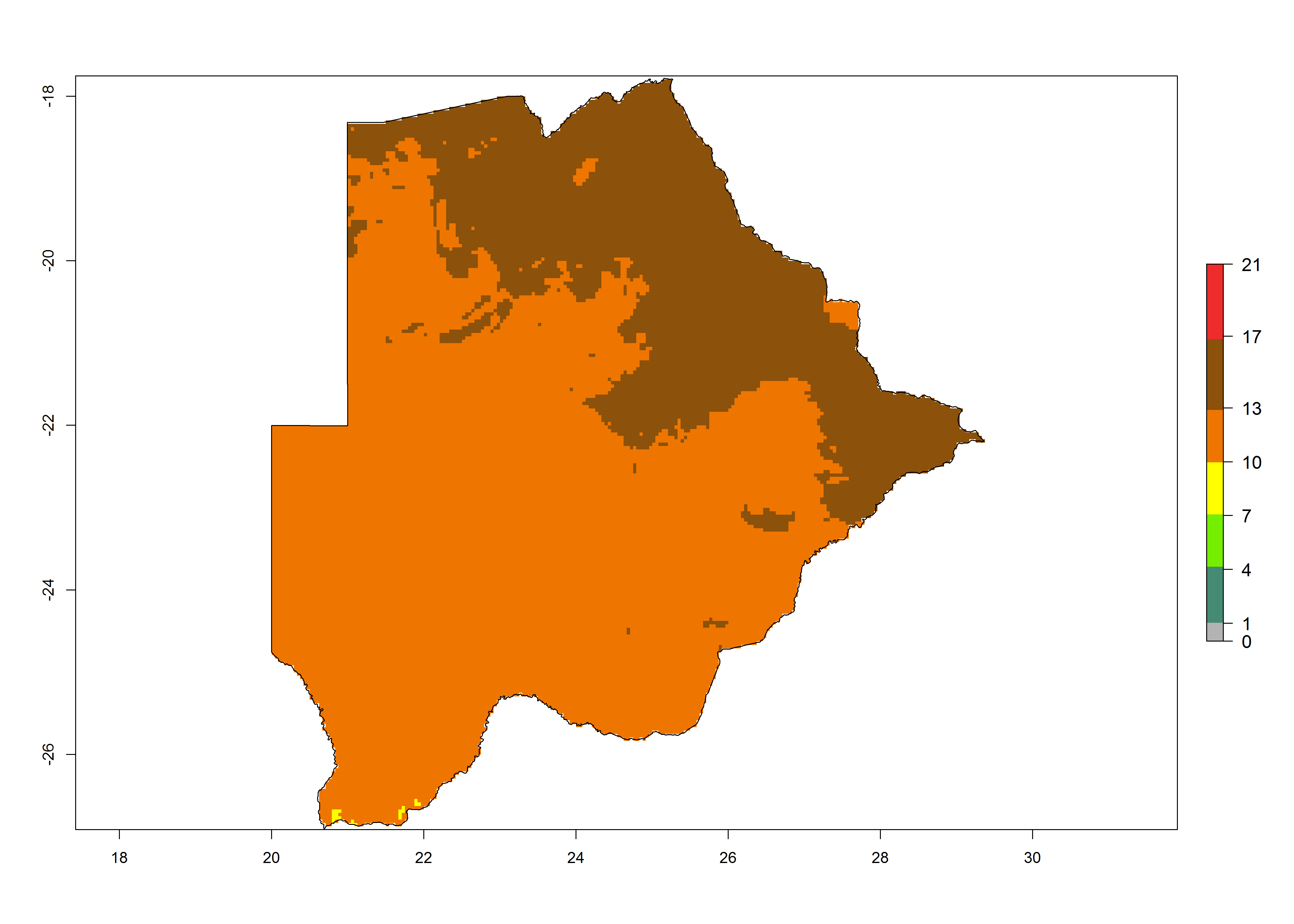
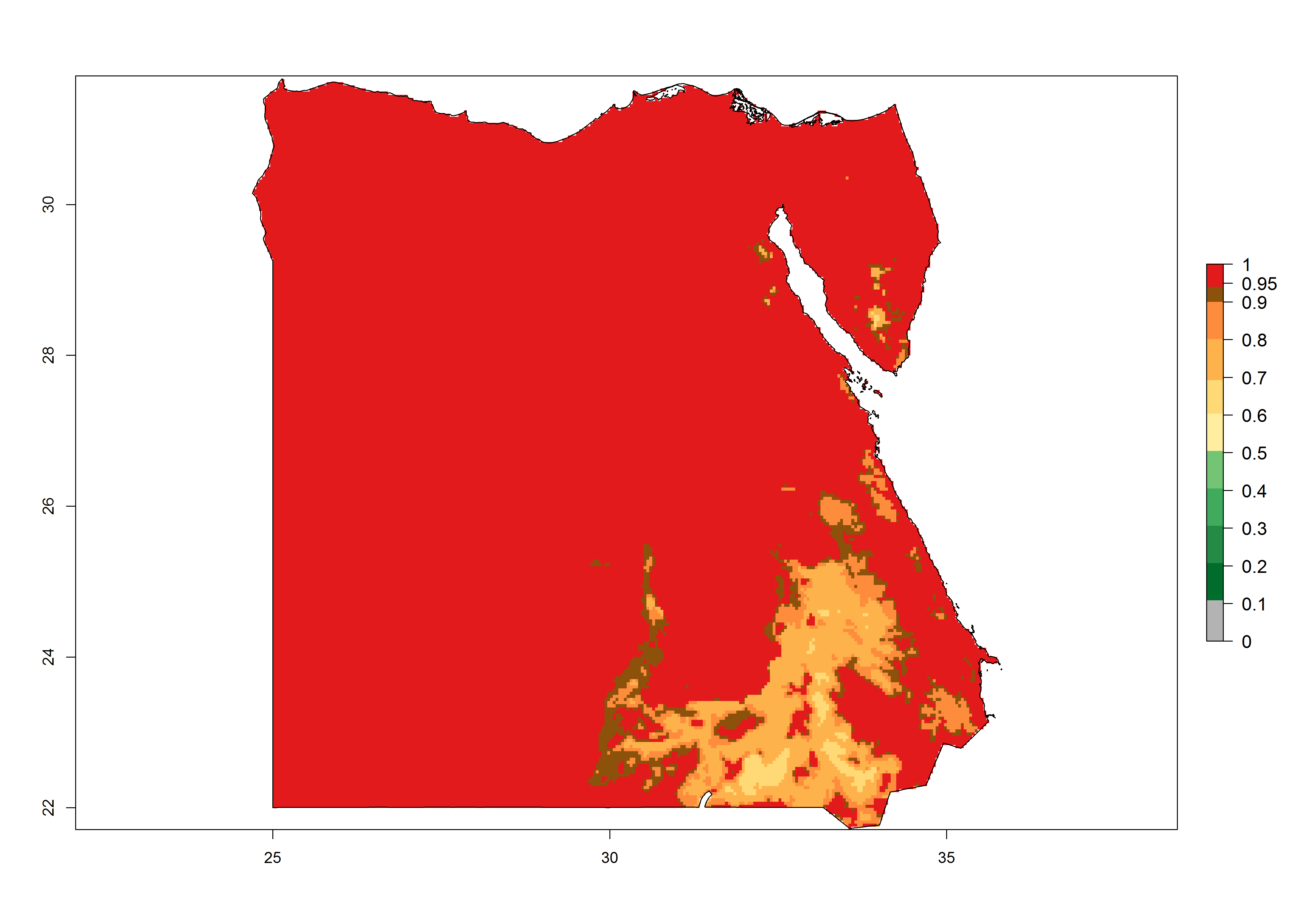
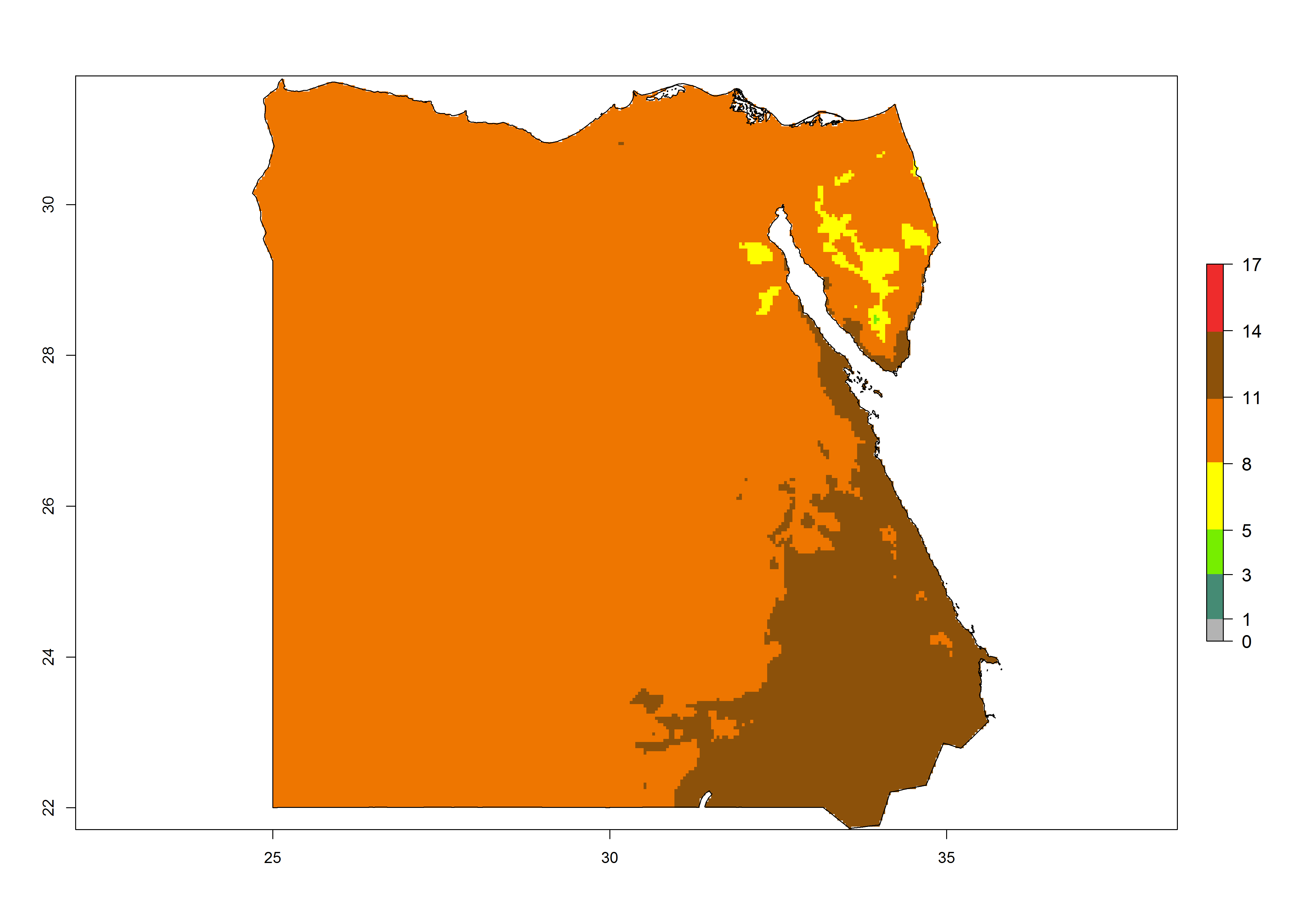
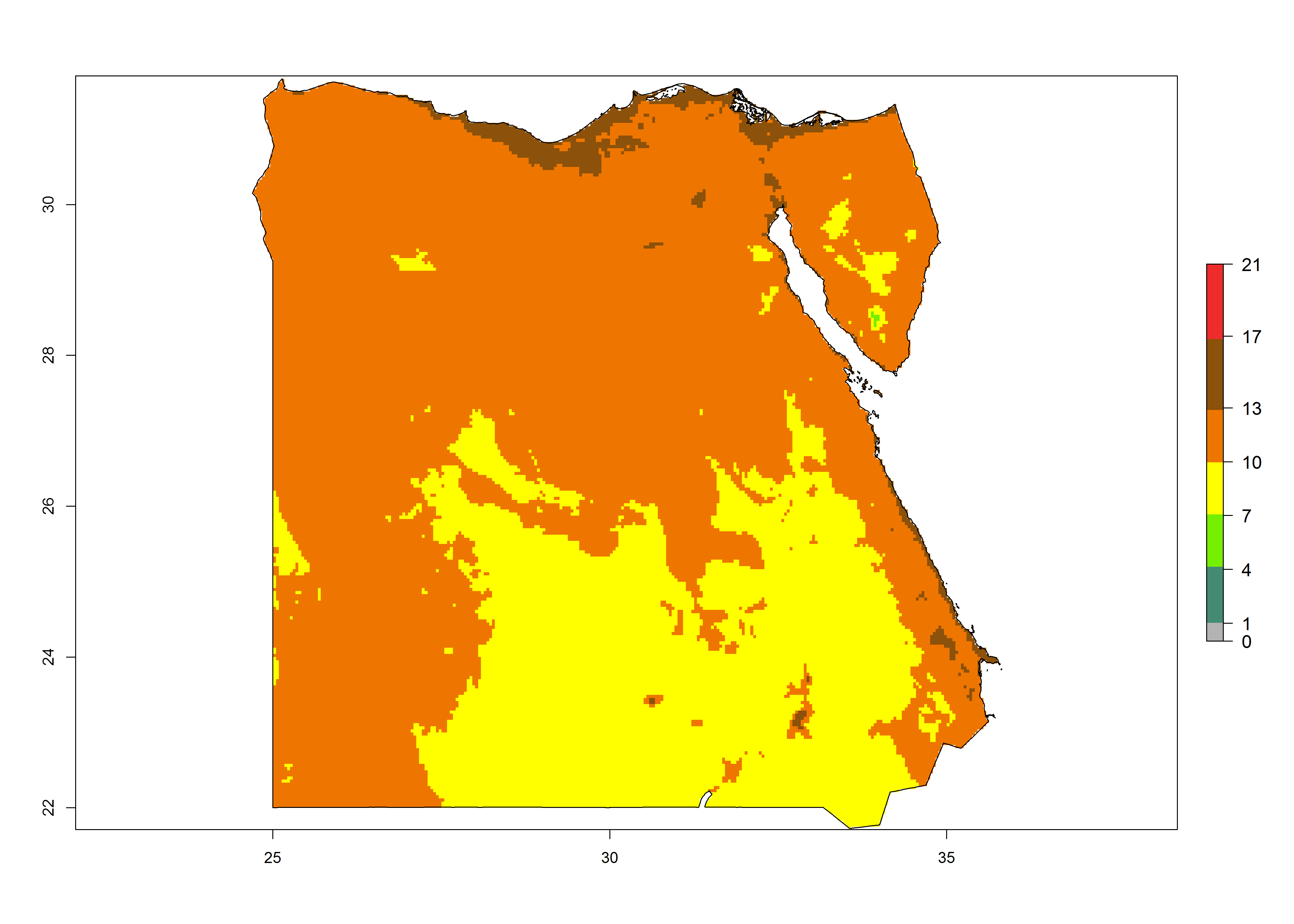
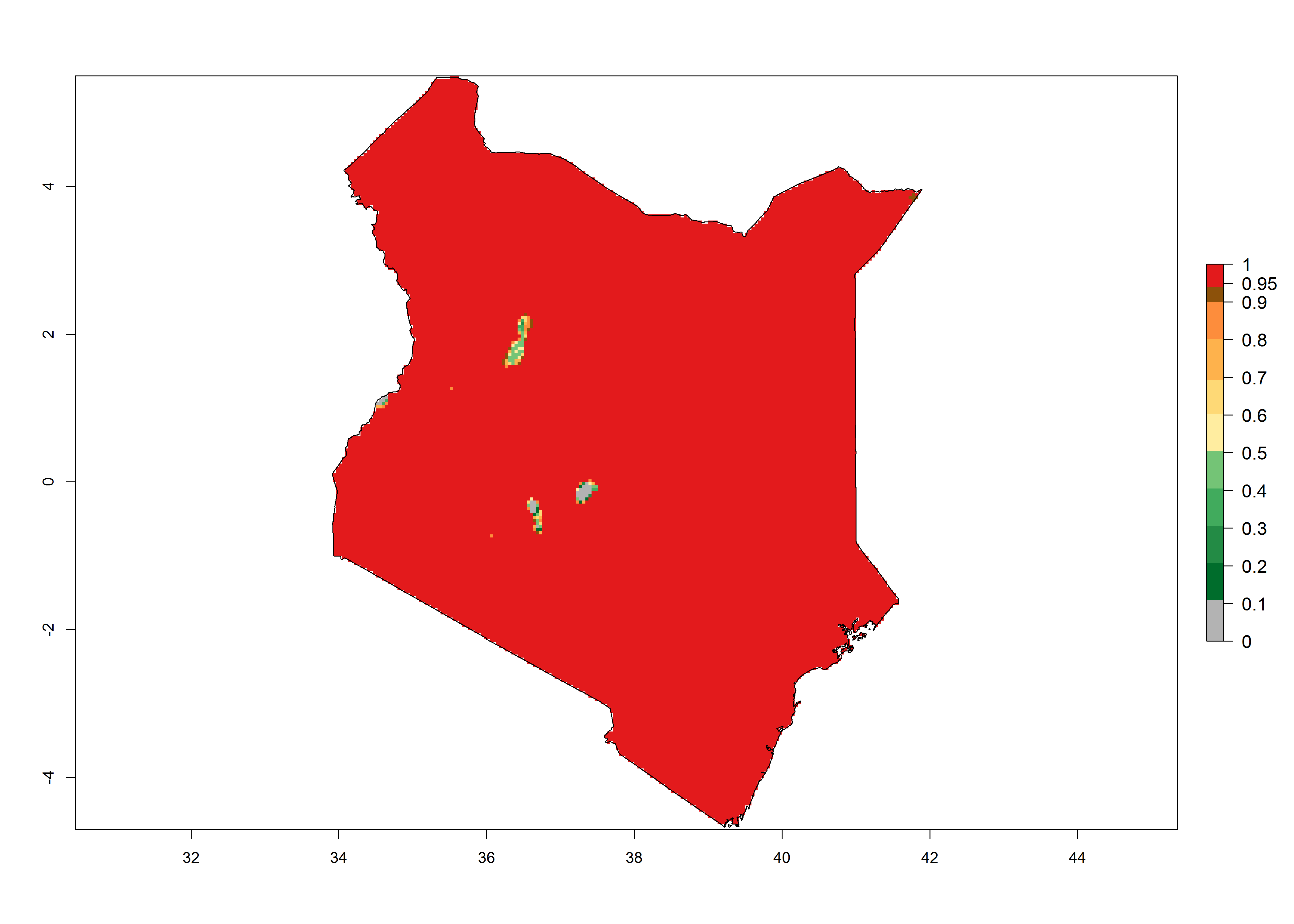
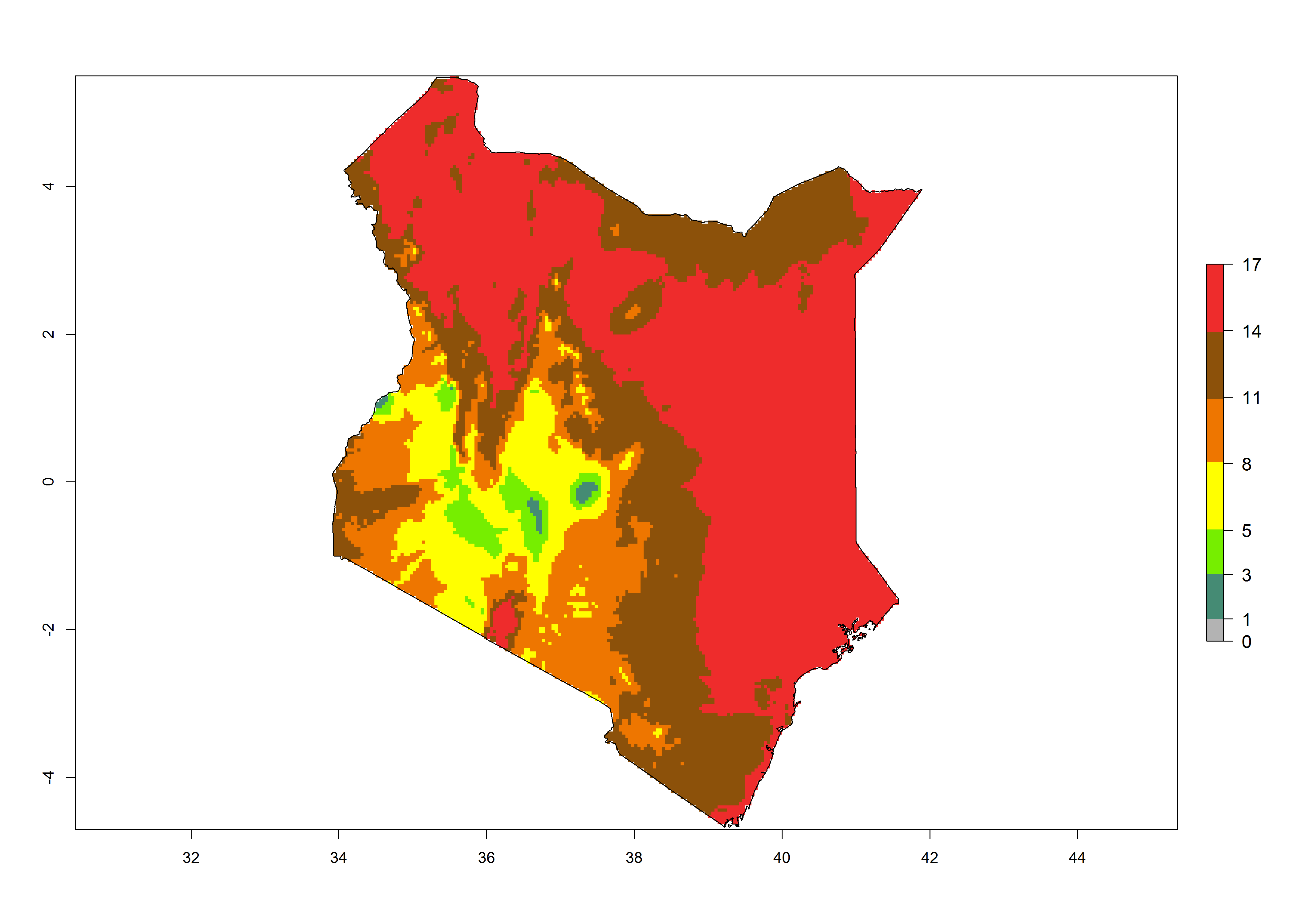
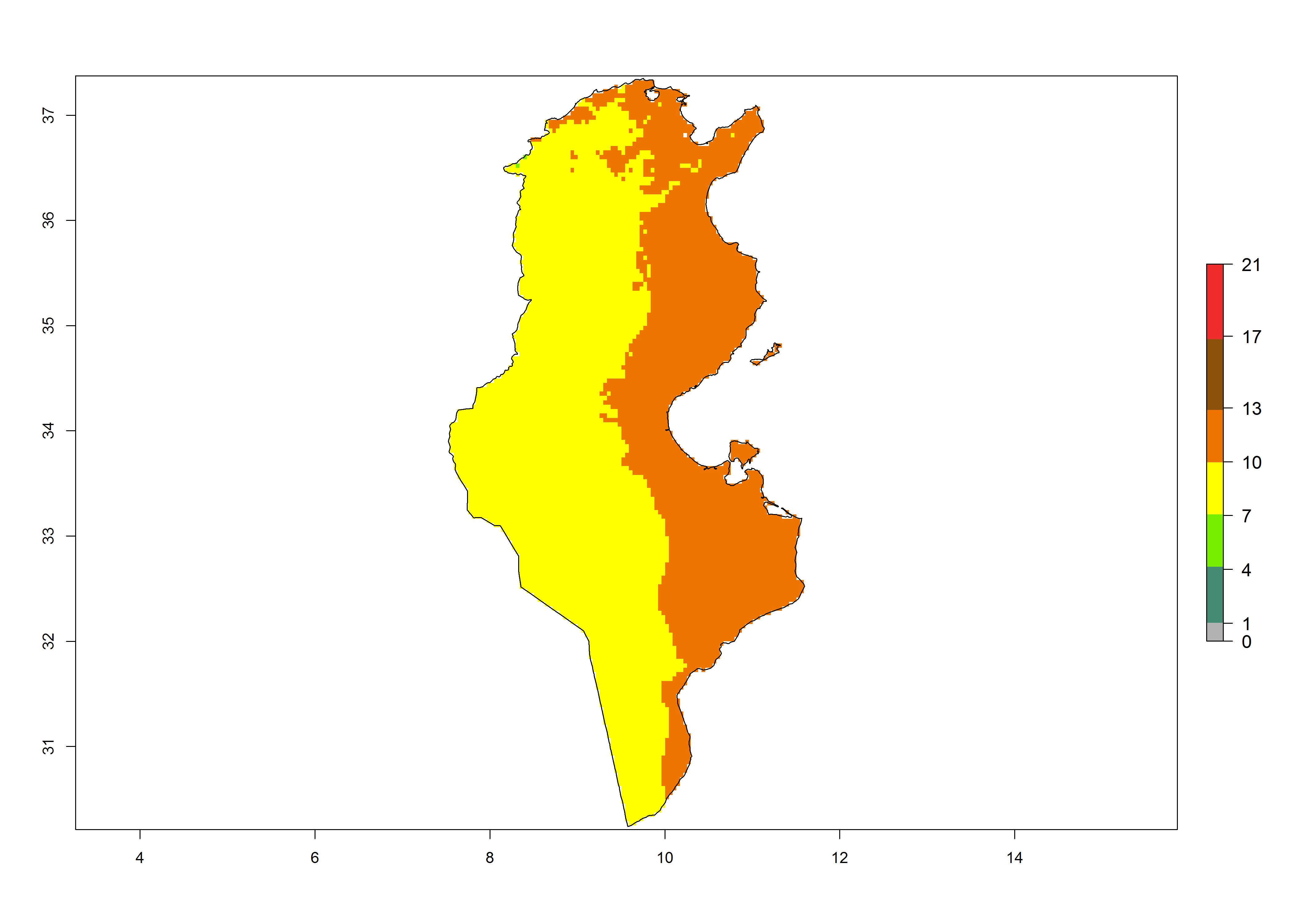
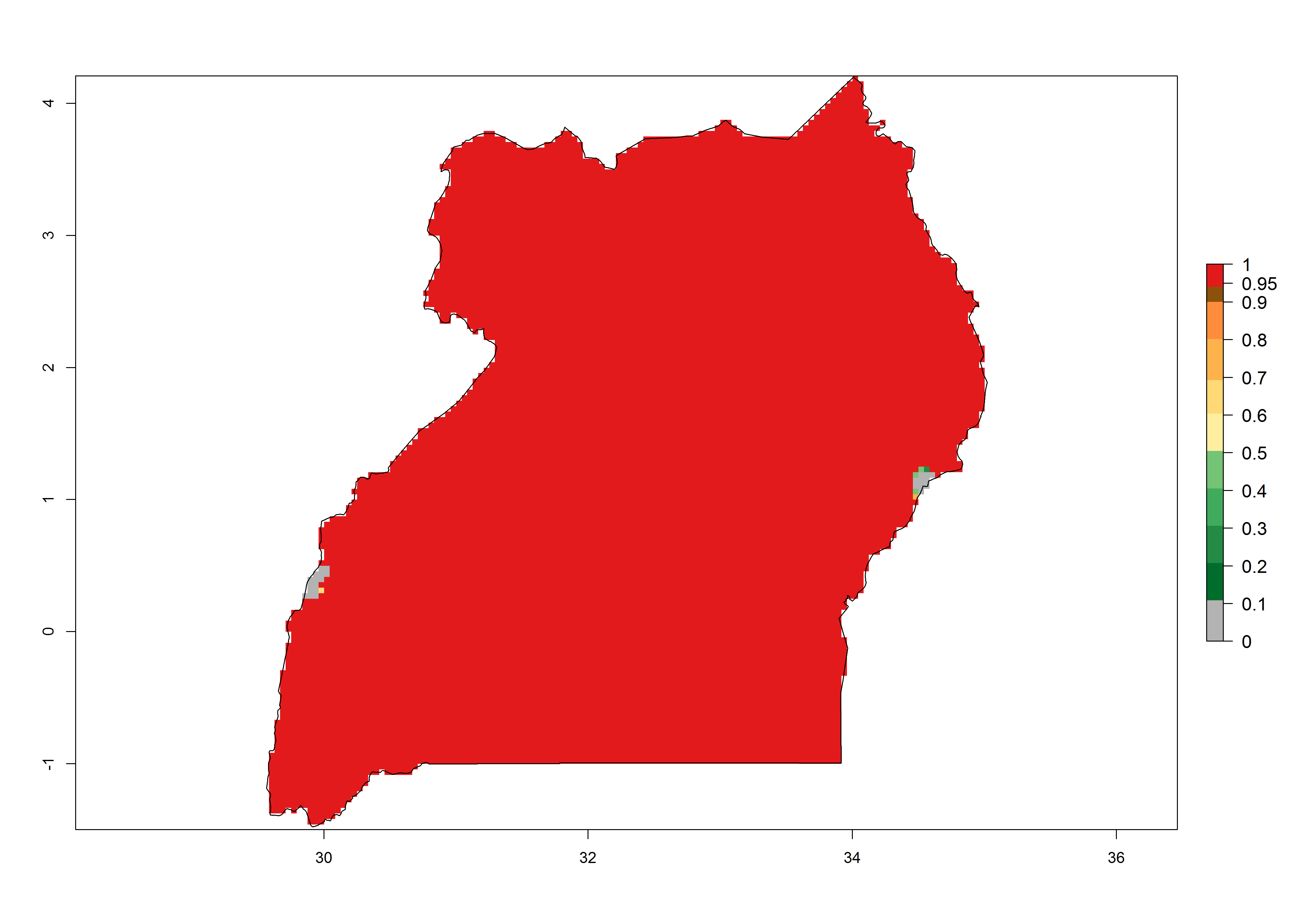
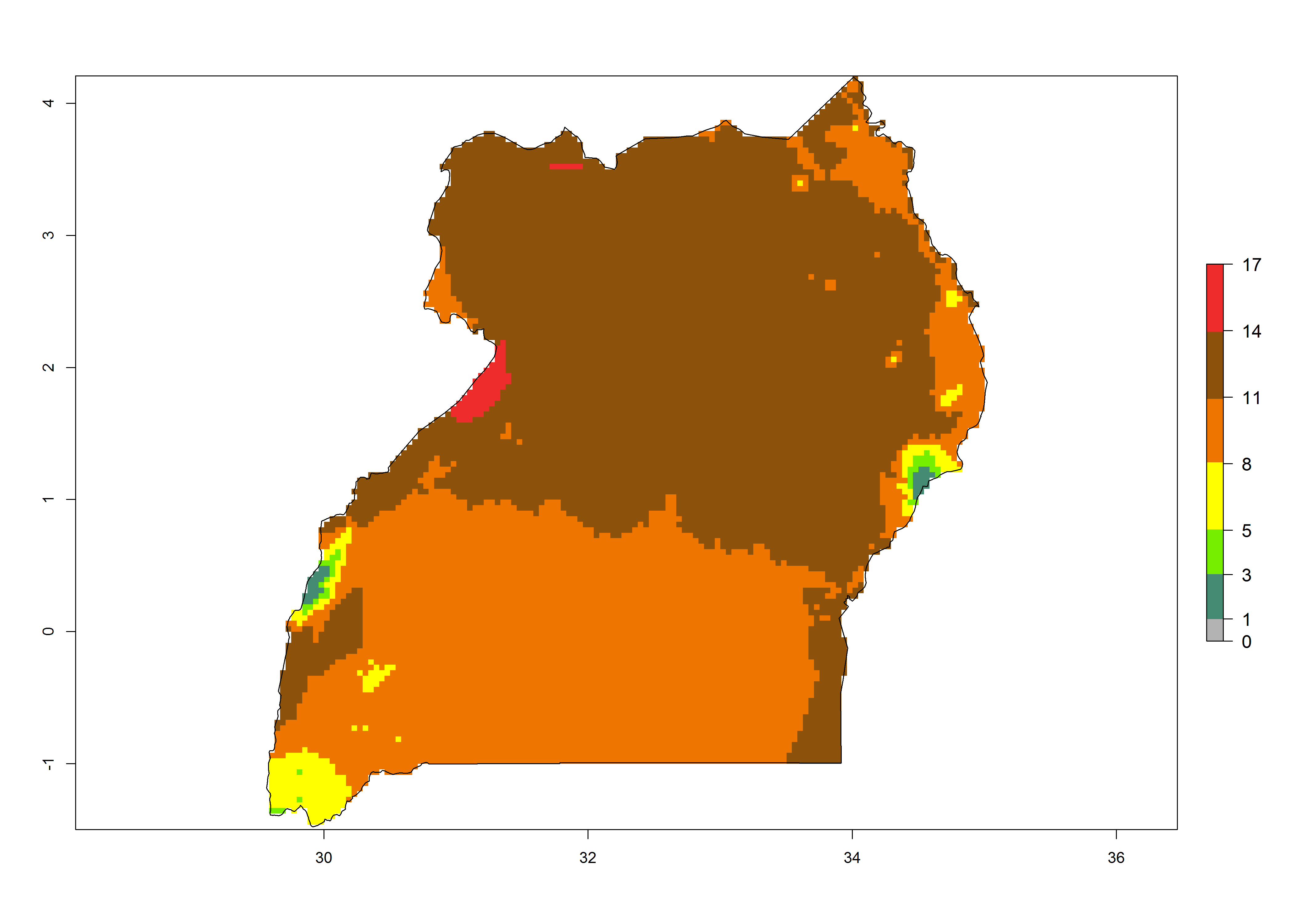
Under the present climate, T. absoluta represents a severe risk (ERI>0.95) in Western and Eastern countries, which is associated with a GI>14–17 generations per year and a high AI (AI>17) (Fig. 6). The highest risks of establishment (ERI>0.95, red zones) are observed in production zones of each country. In Mediterranean countries (e.g., Egypt, Morocco and Tunisia), the risk of establishment is high but GI gives values between 10-13 (Egypt) and 5-8 (Morocco) generations per year and with an AI<13 for both countries.
| ERI | GI | AI |
| a) Botswana | ||
 |
 |
 |
| b) Egypt | ||
 |
 |
 |
| c) Kenya | ||
 |
 |
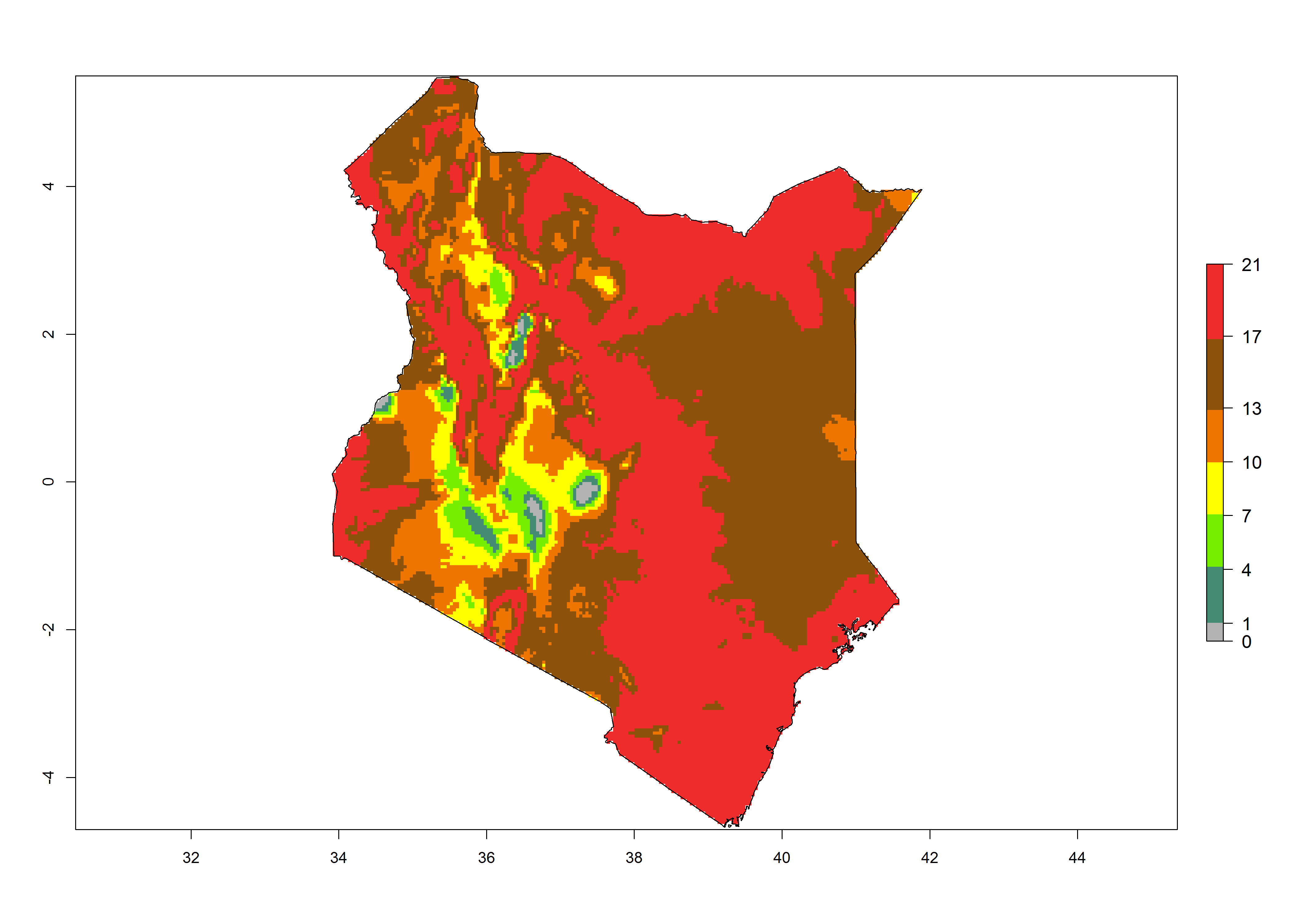 |
| d) Morocco | ||
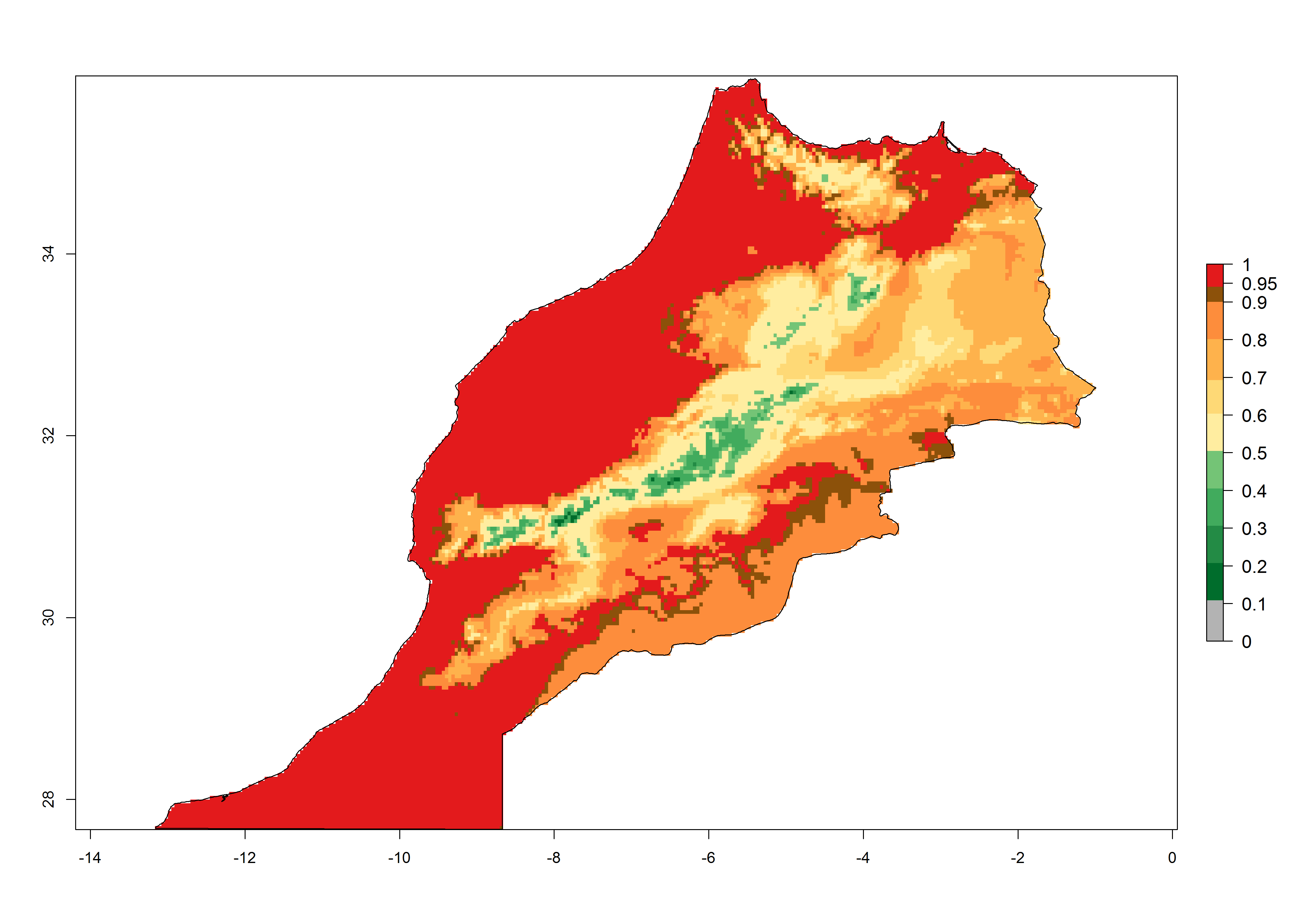 |
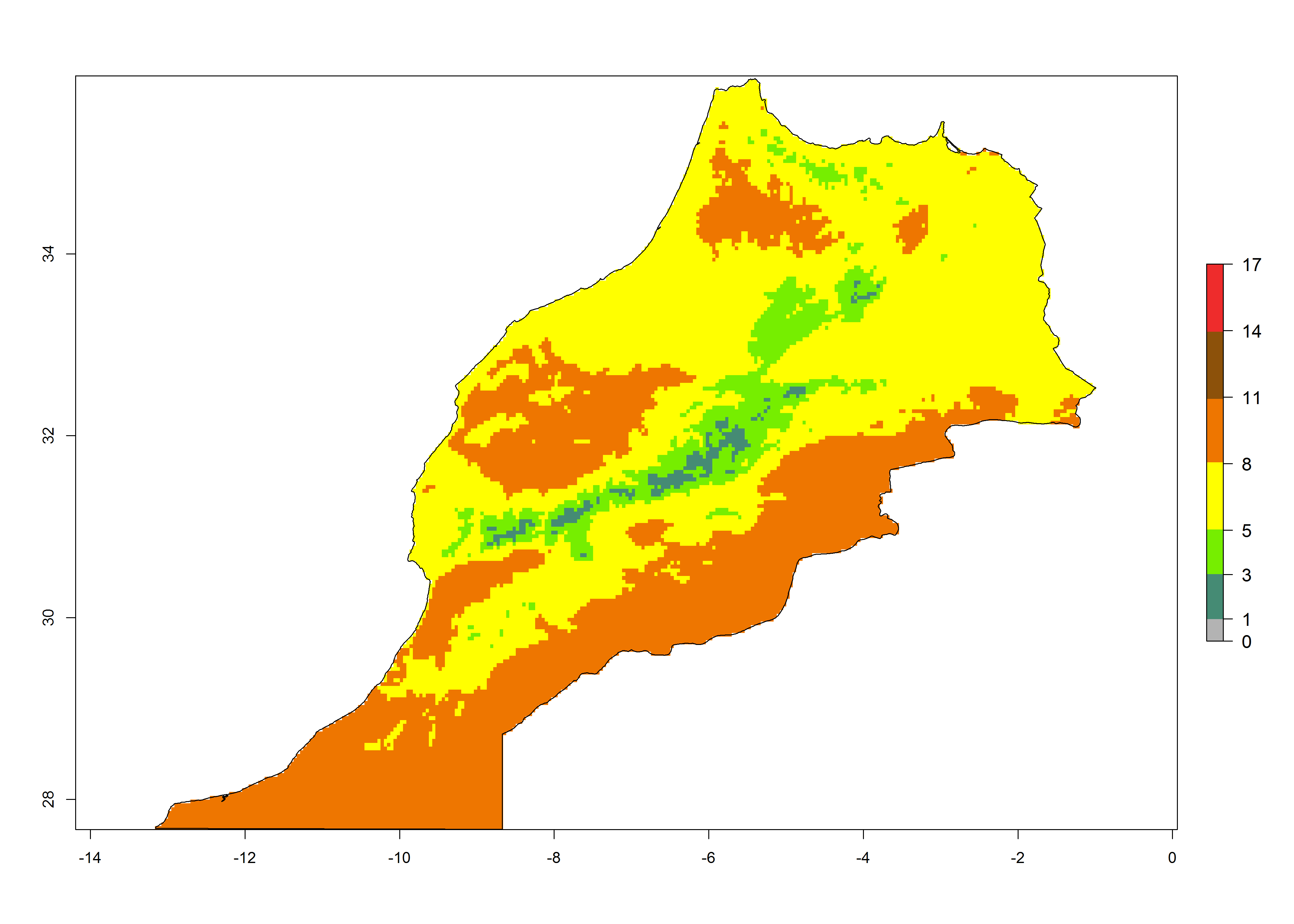 |
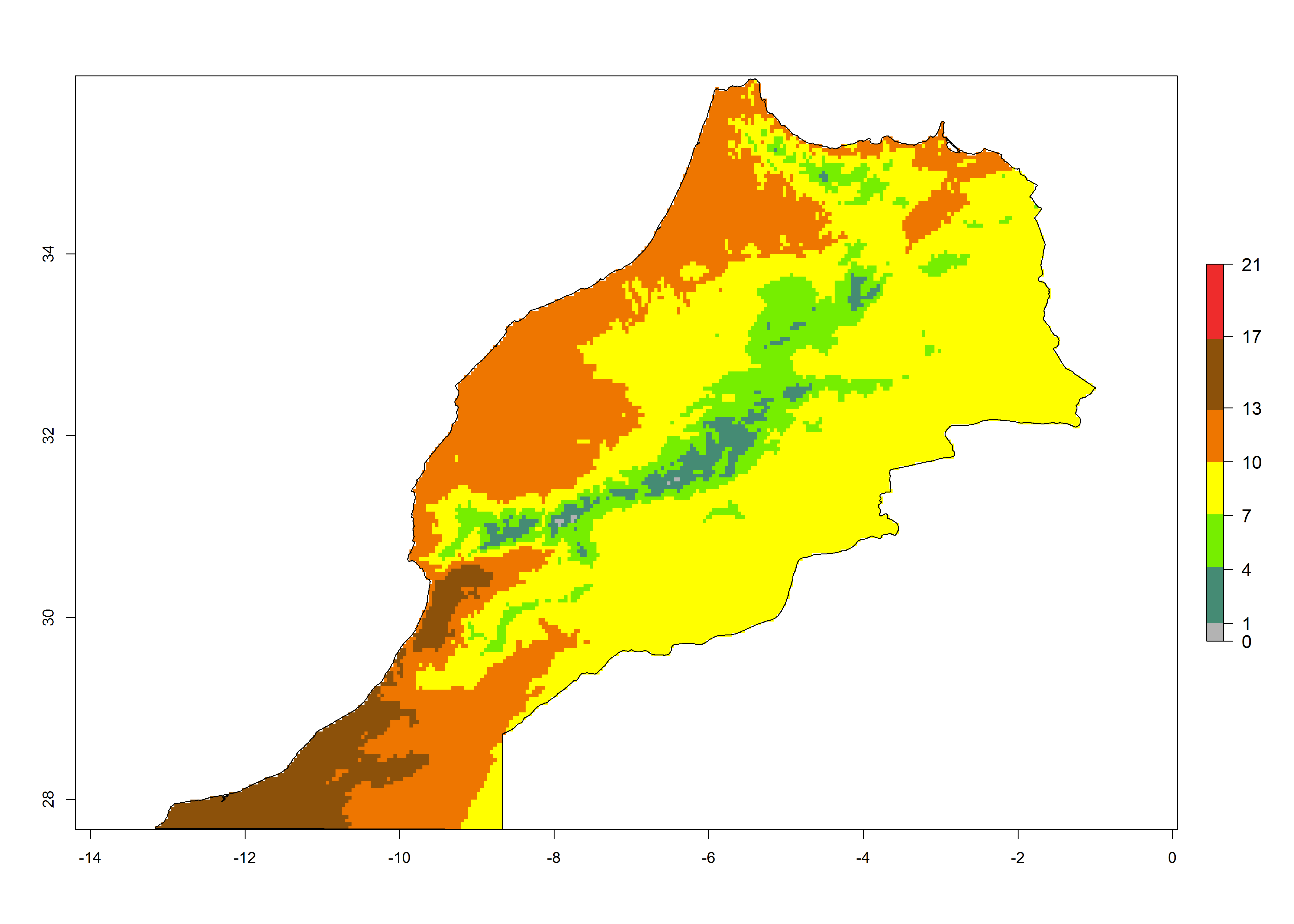 |
| e) Namibia | ||
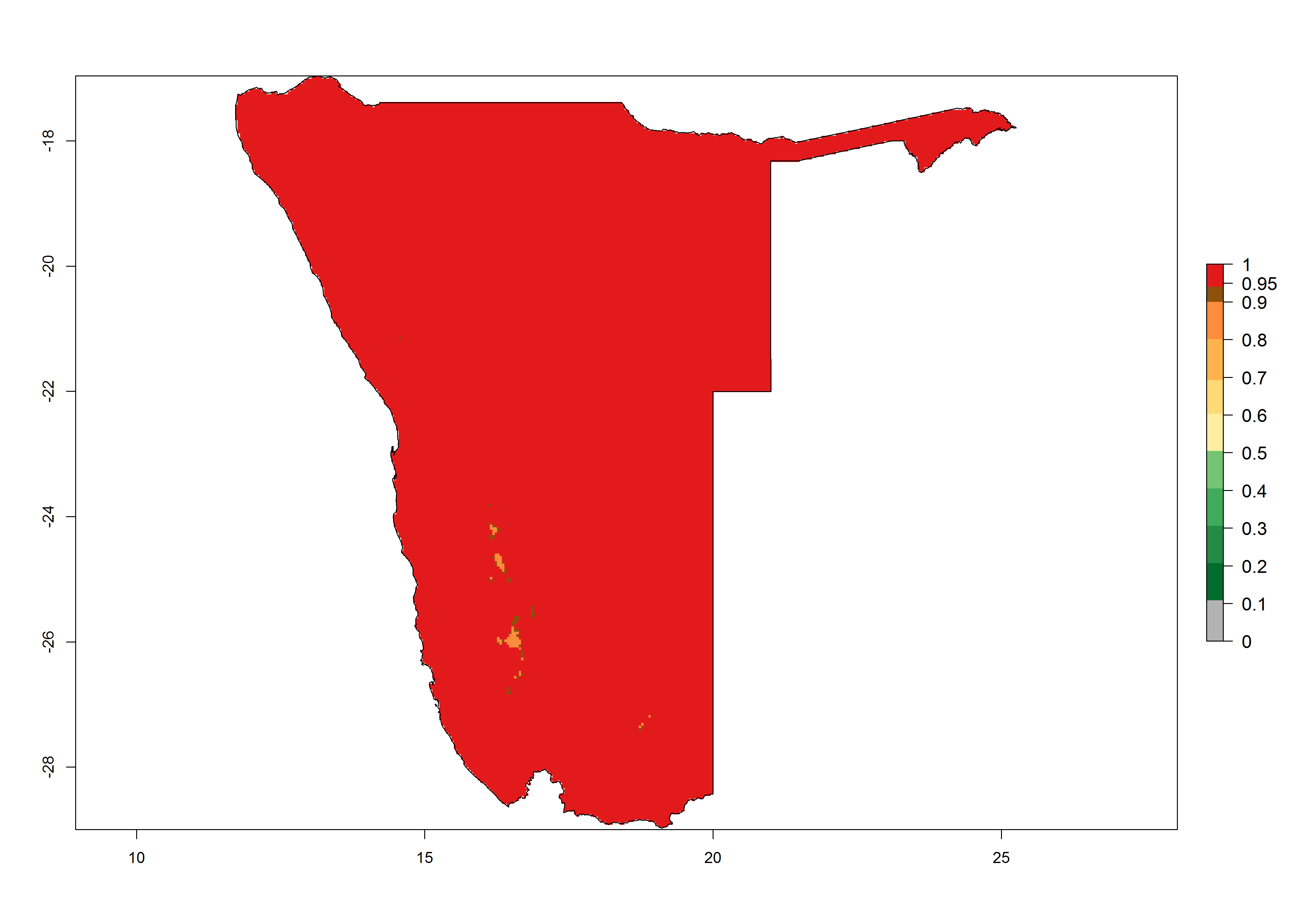 |
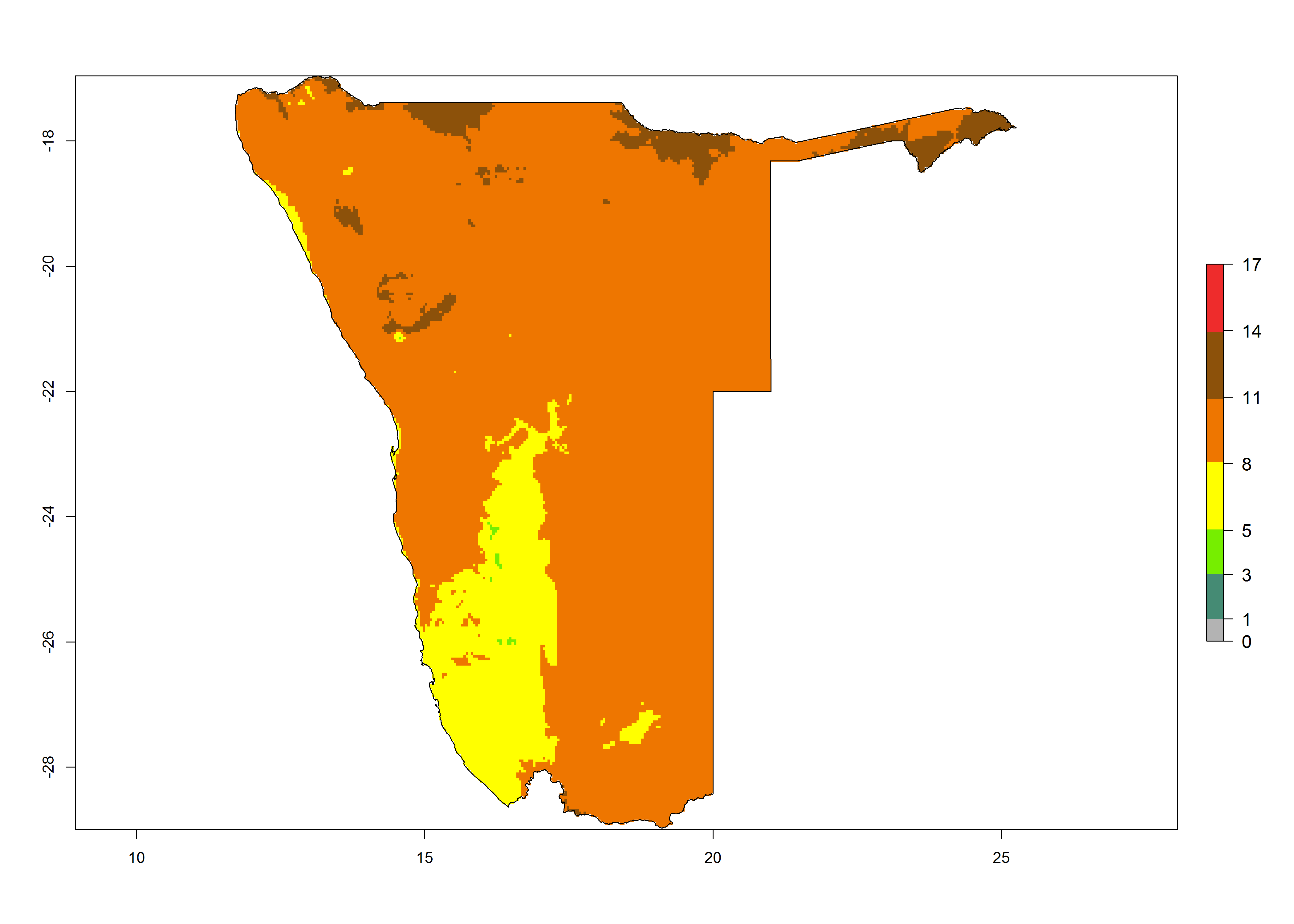 |
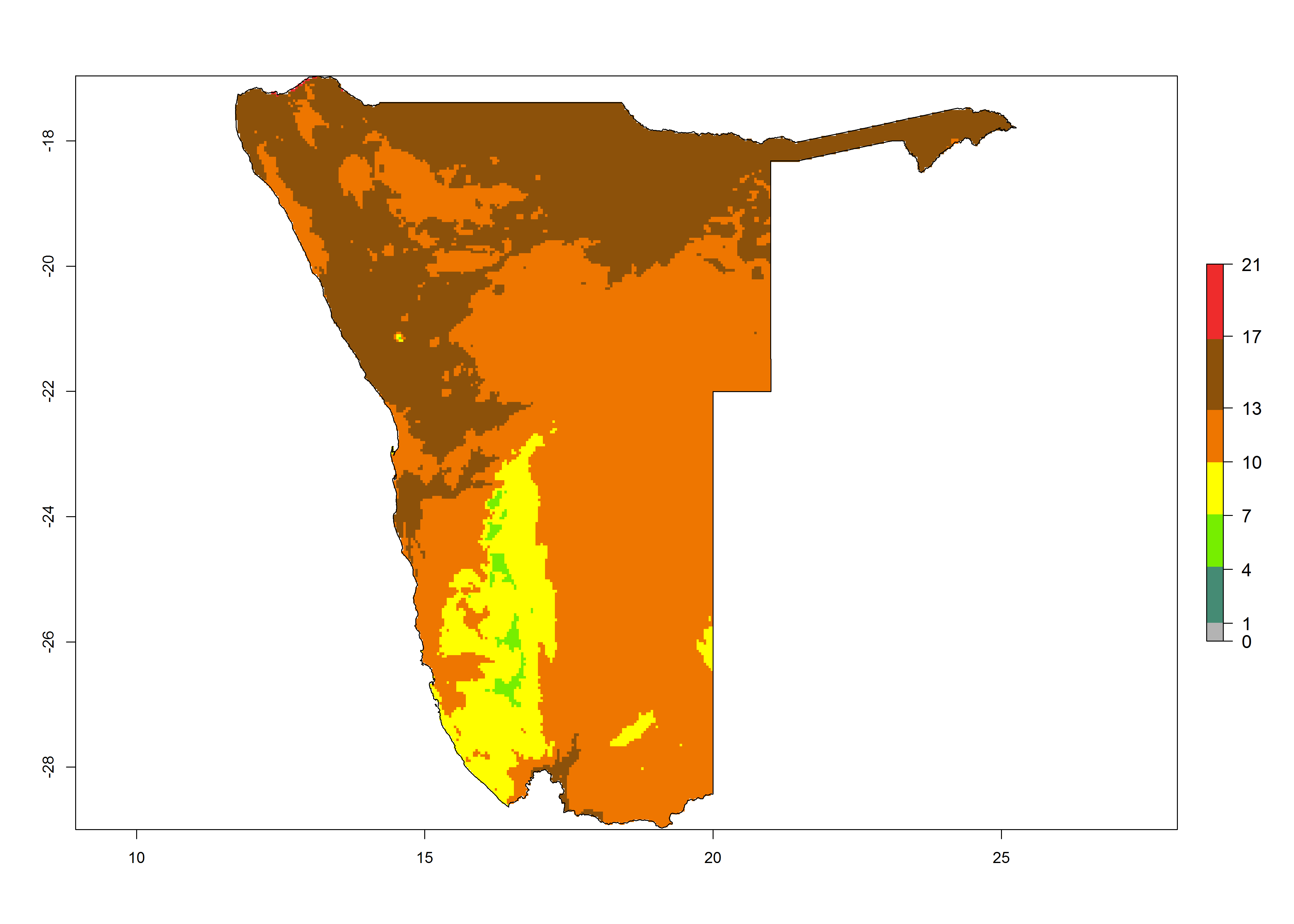 |
| f) Niger | ||
 |
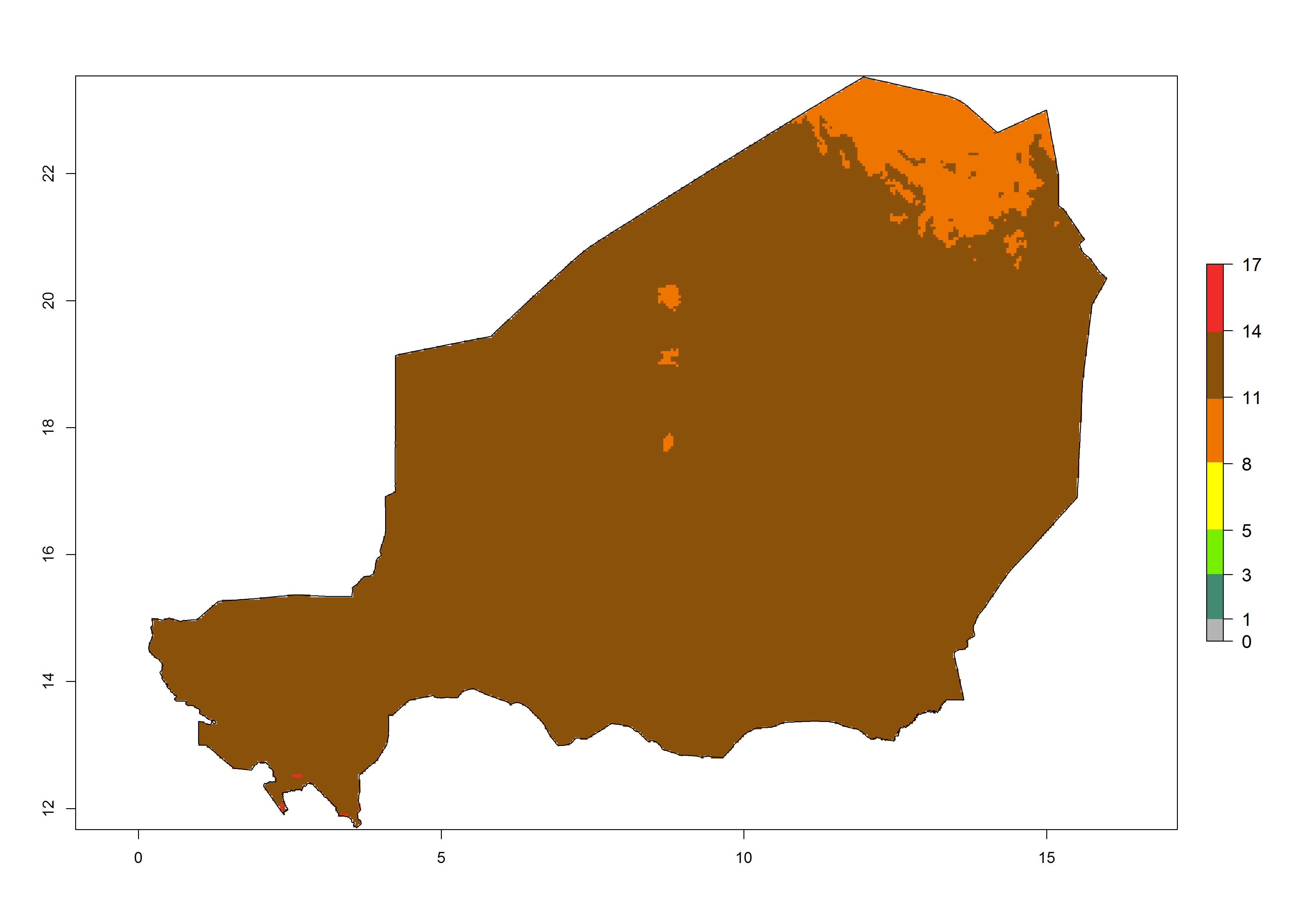 |
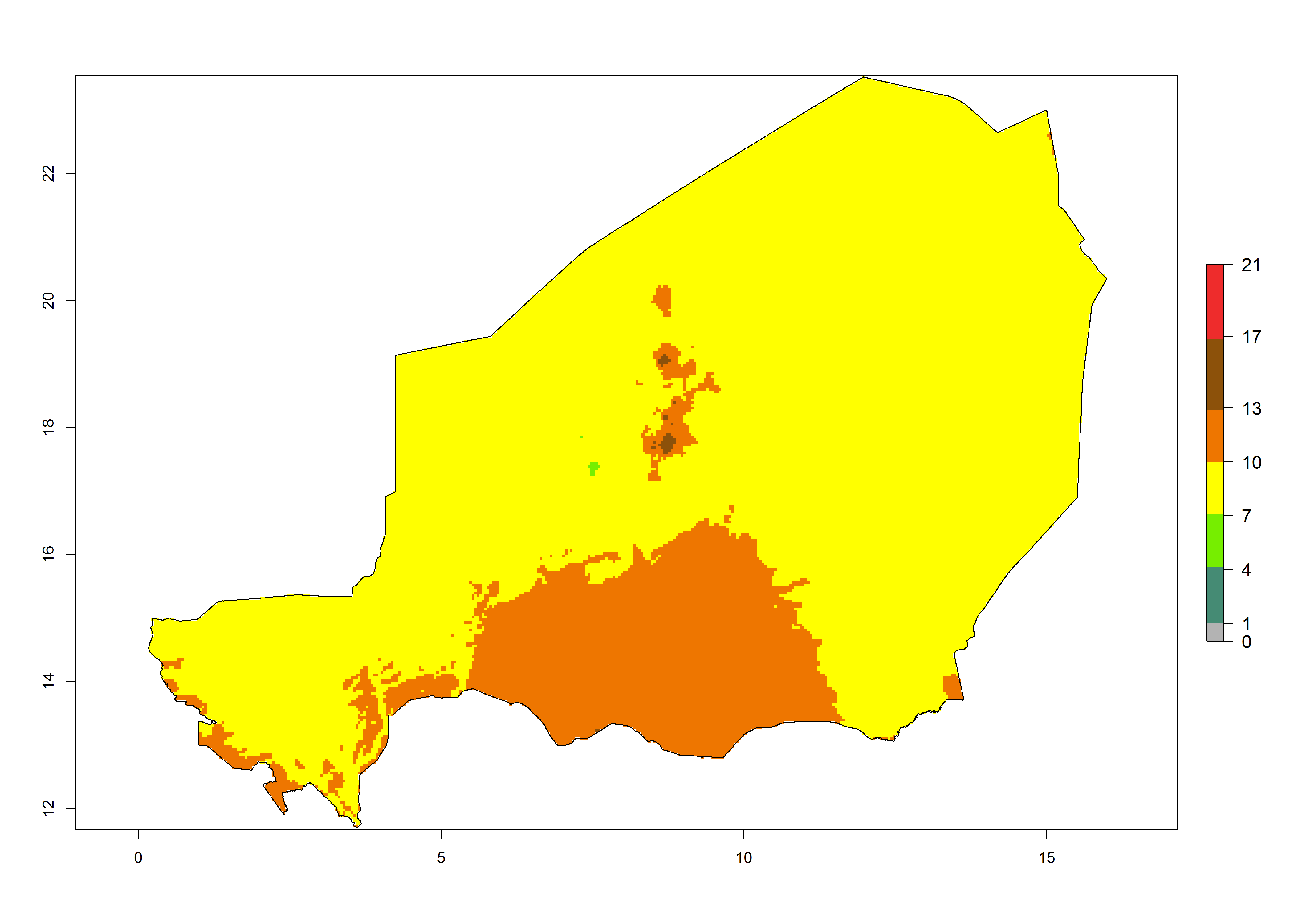 |
| g) Nigeria | ||
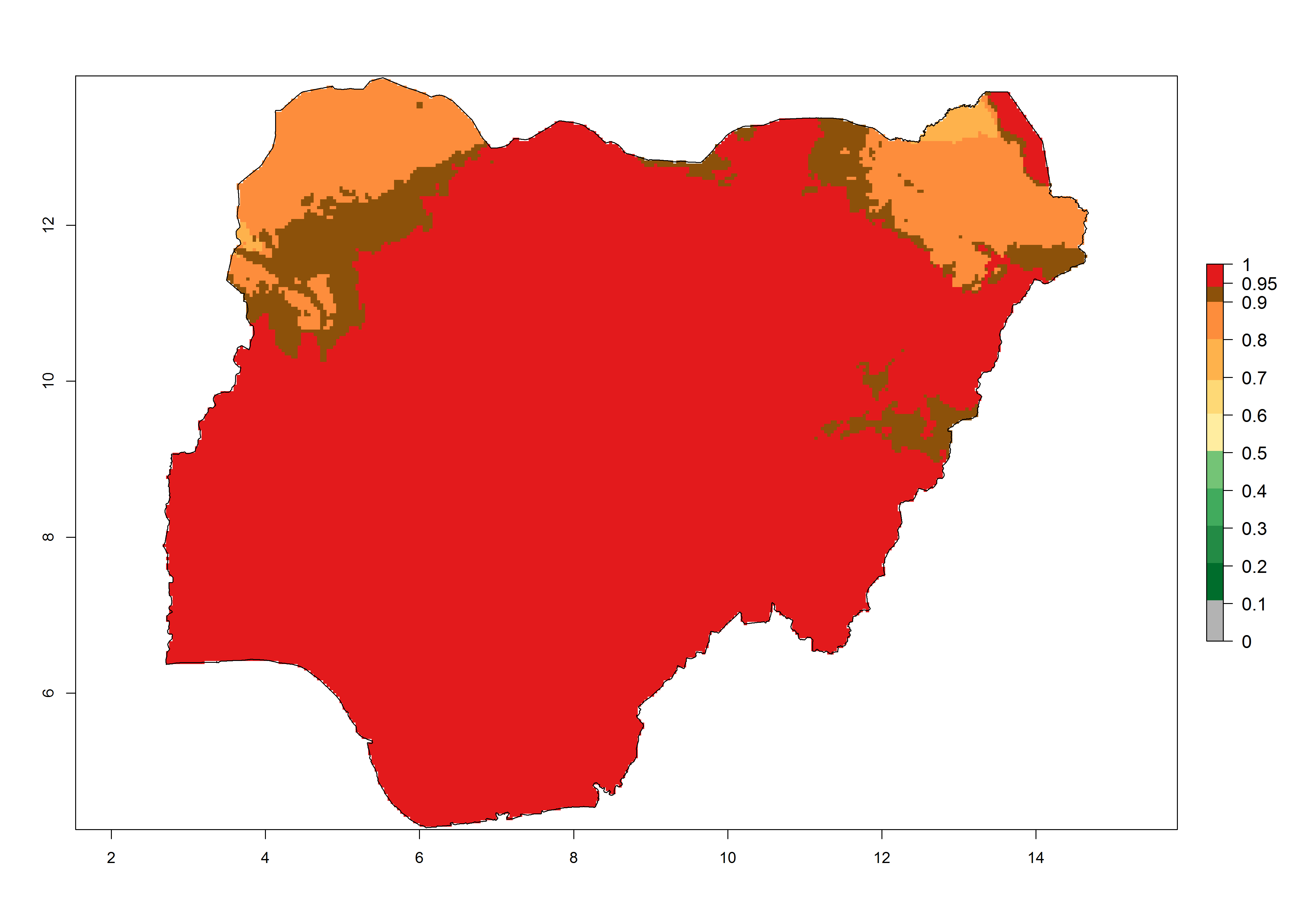 |
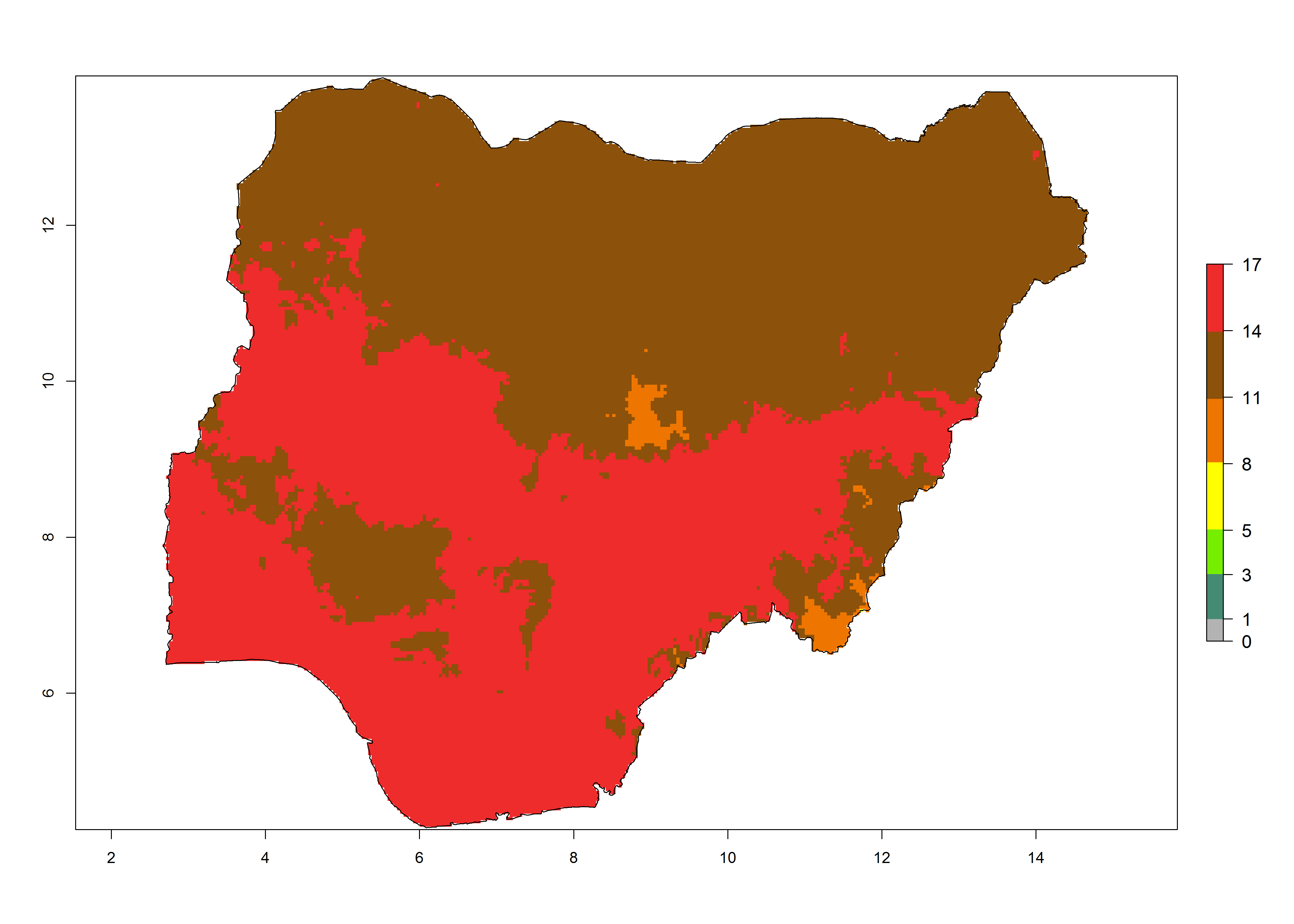 |
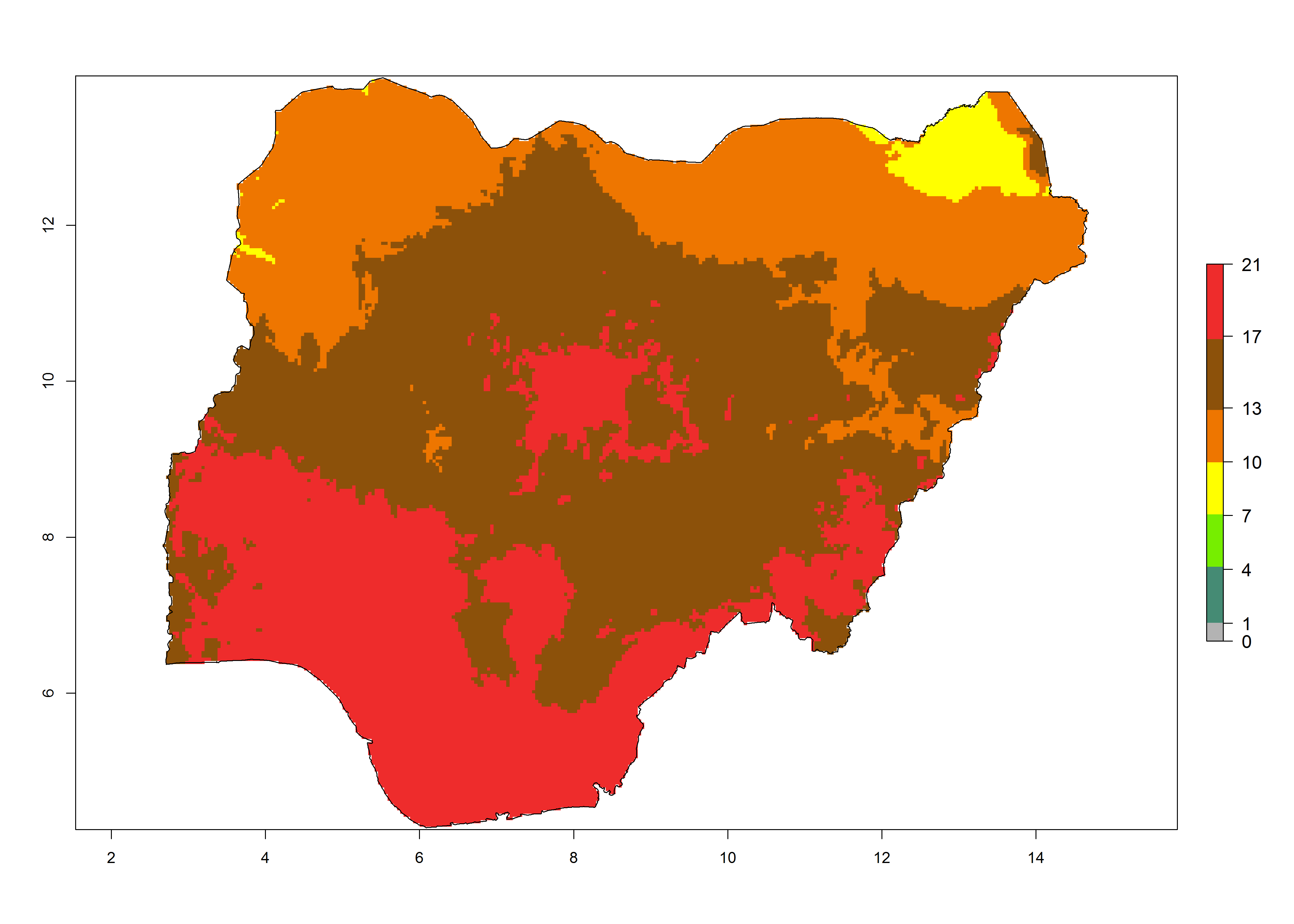 |
| h) Rwanda | ||
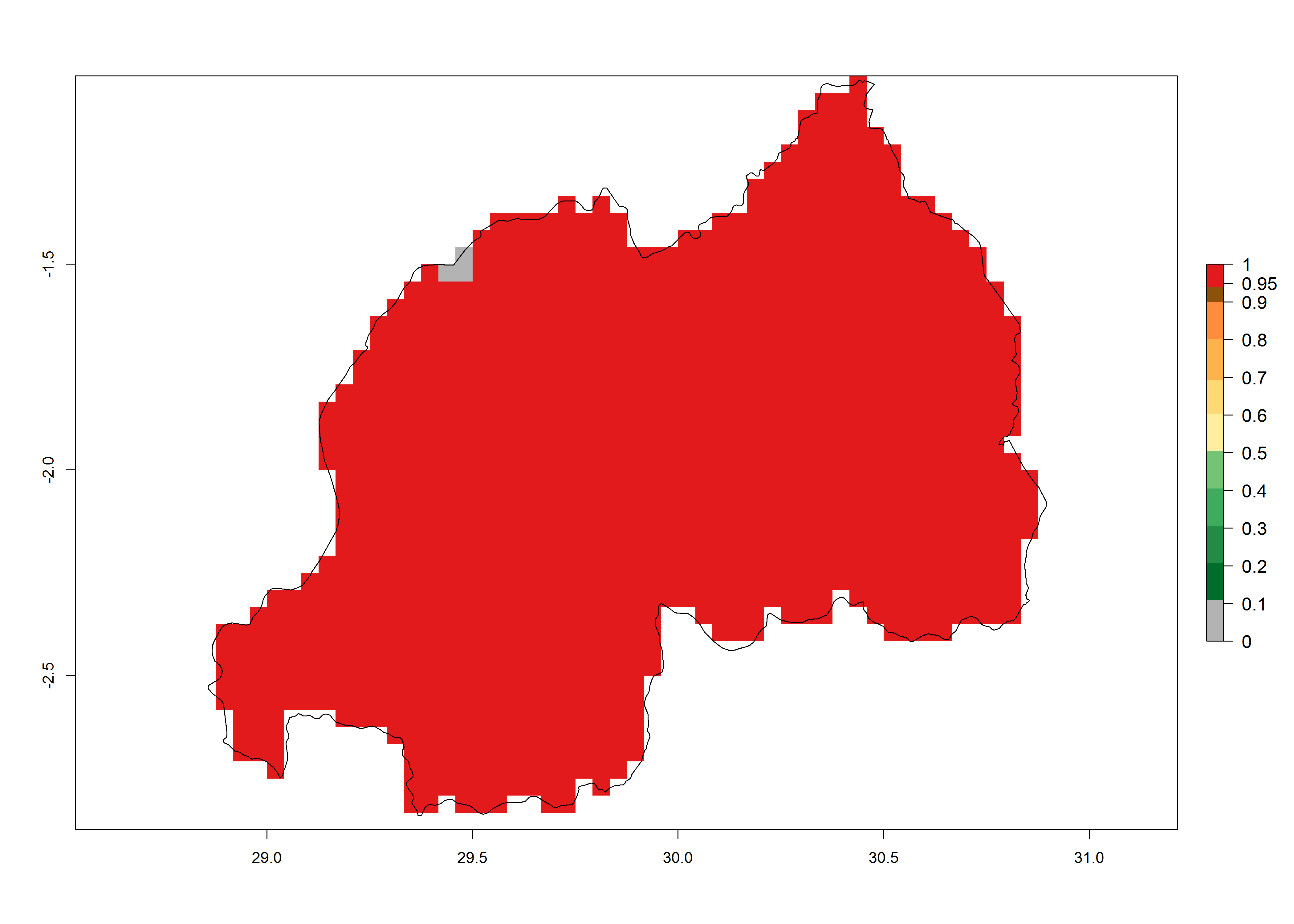 |
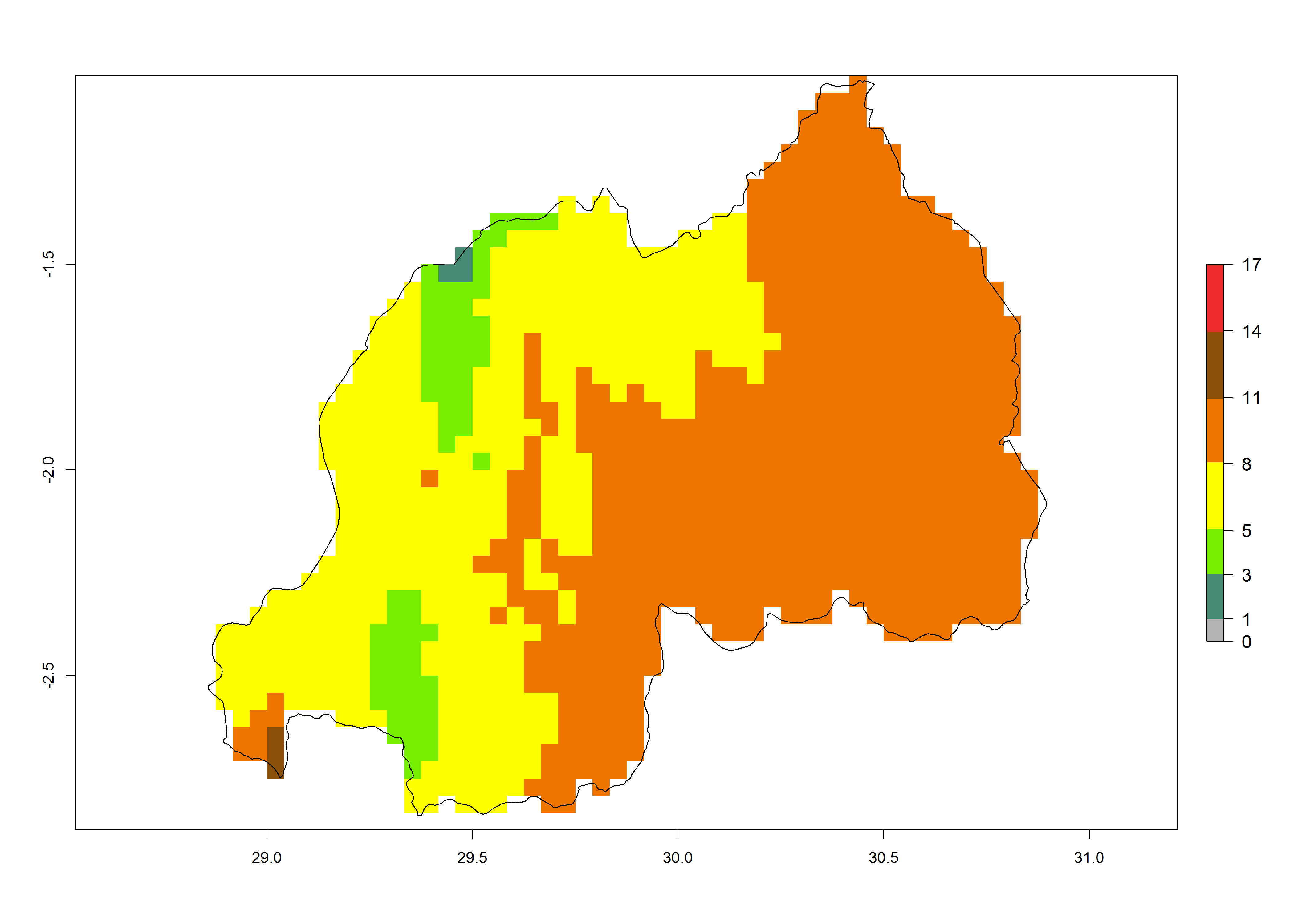 |
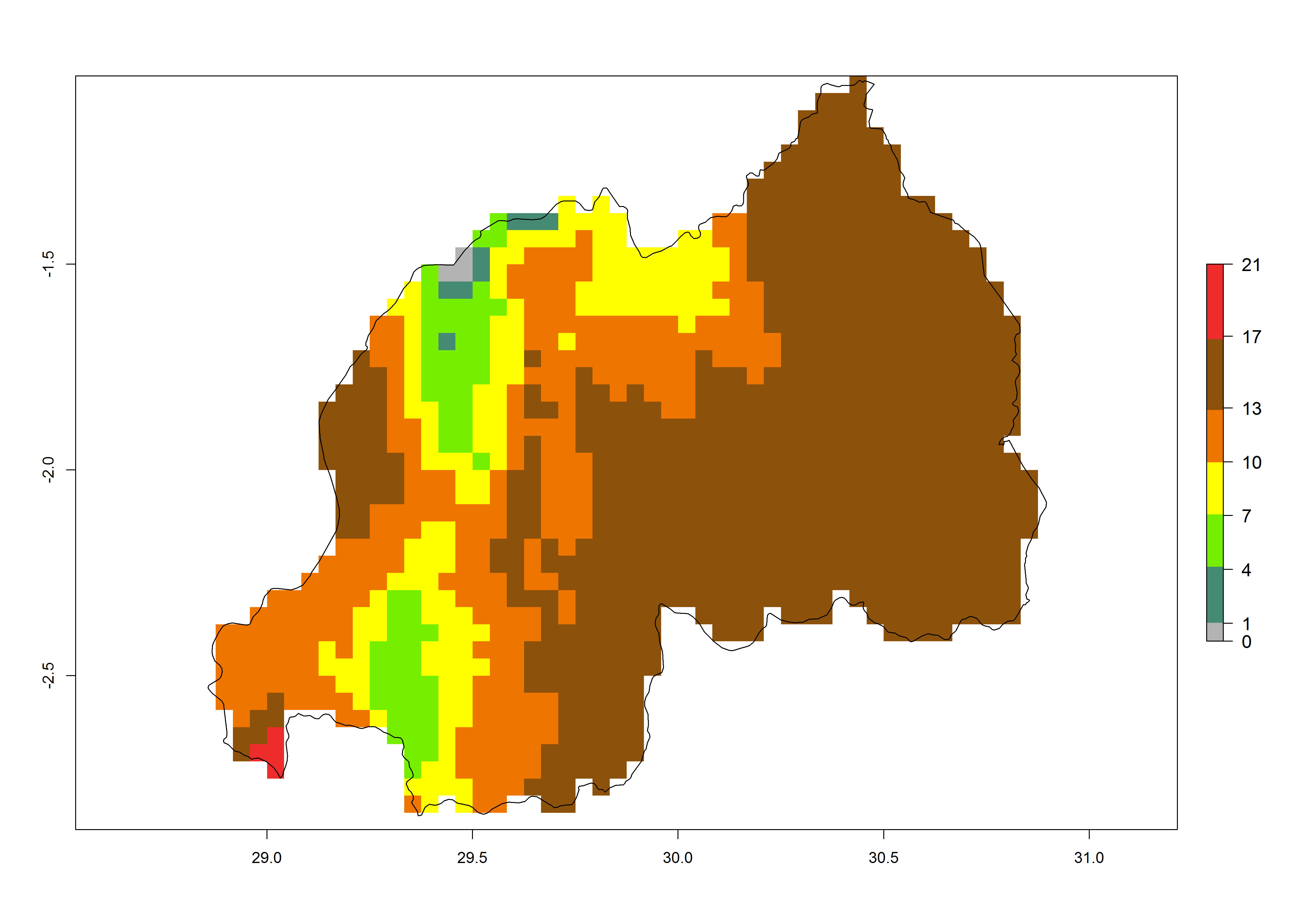 |
| i) South Africa | ||
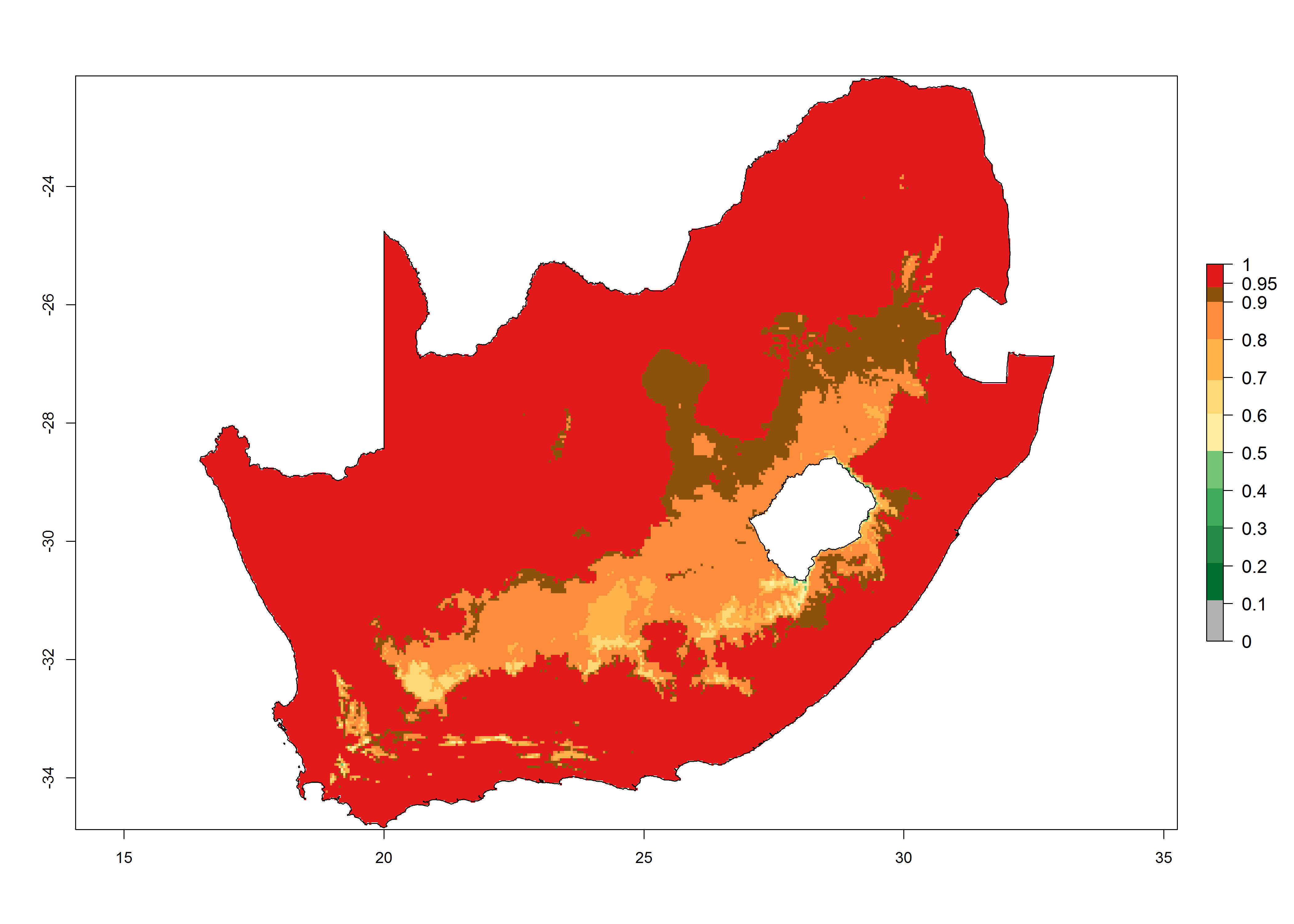 |
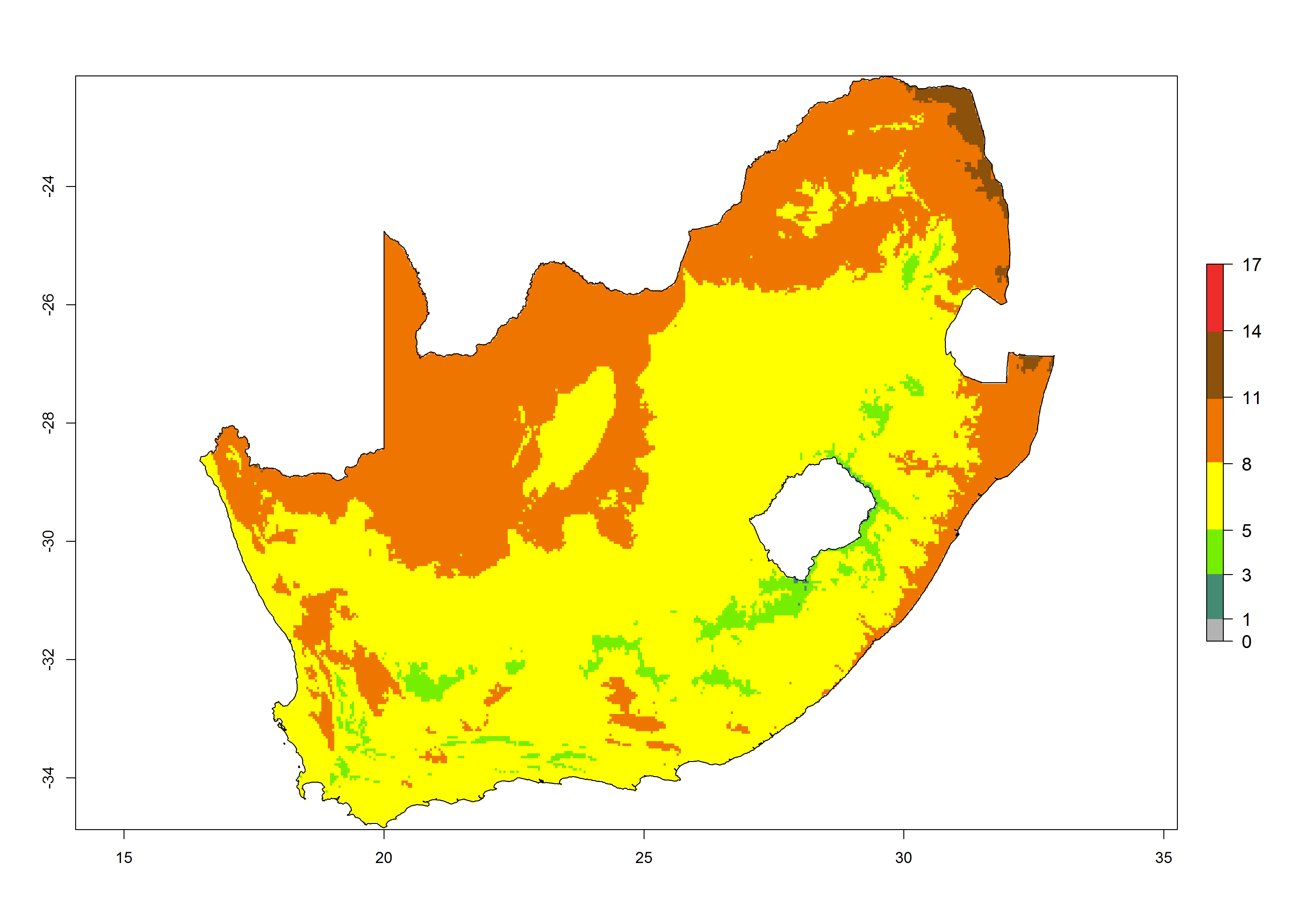 |
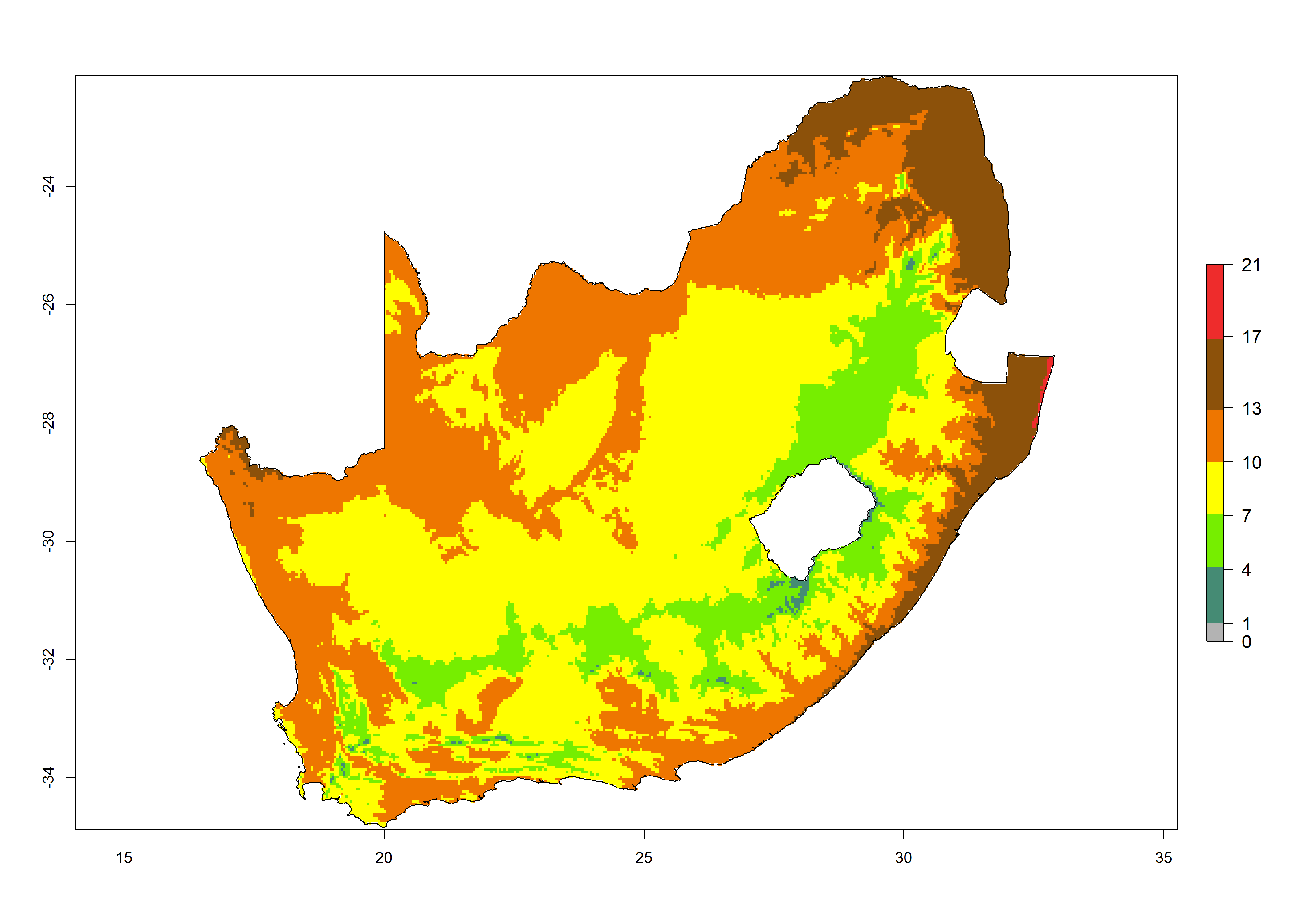 |
| j) Tanzania | ||
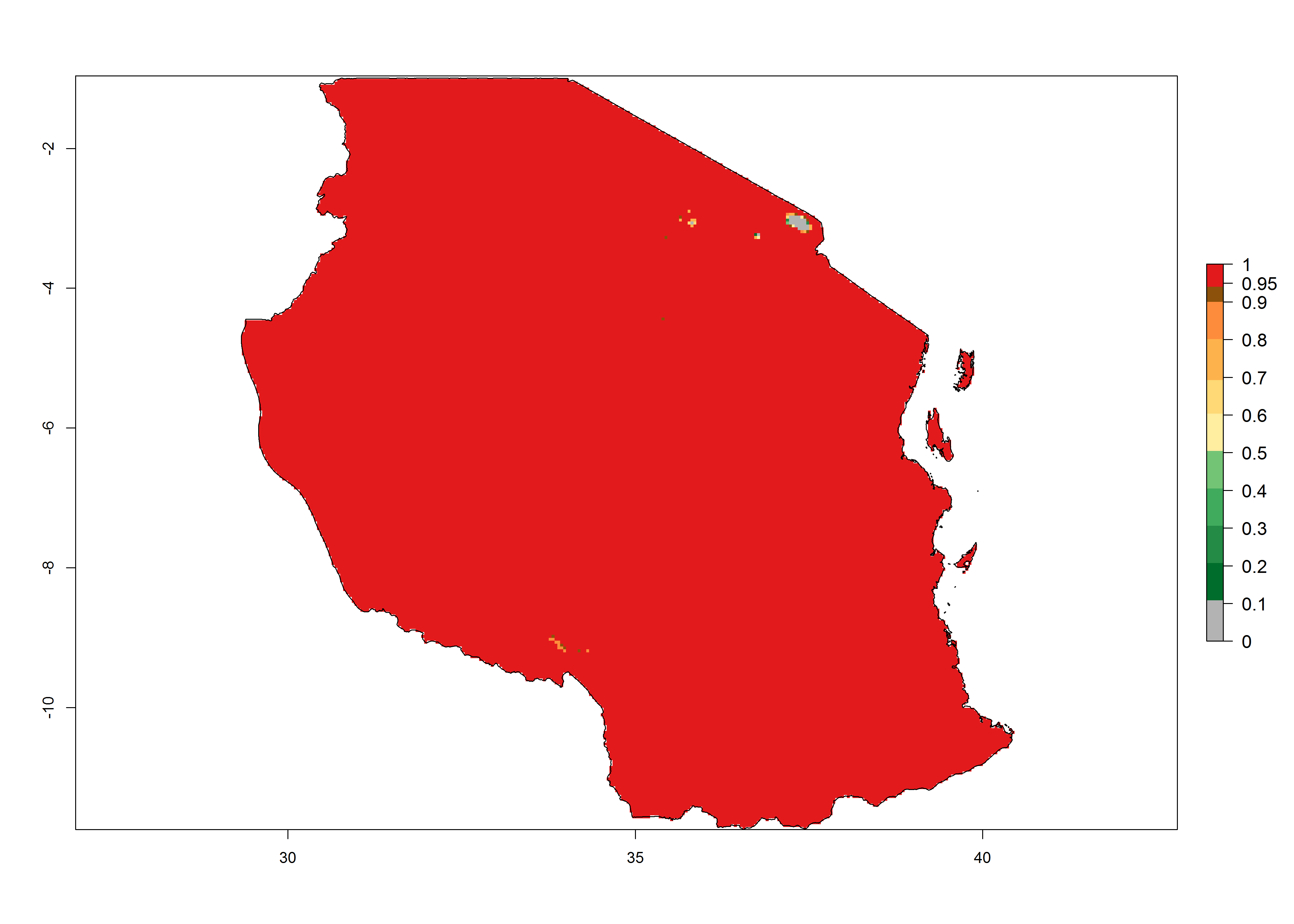 |
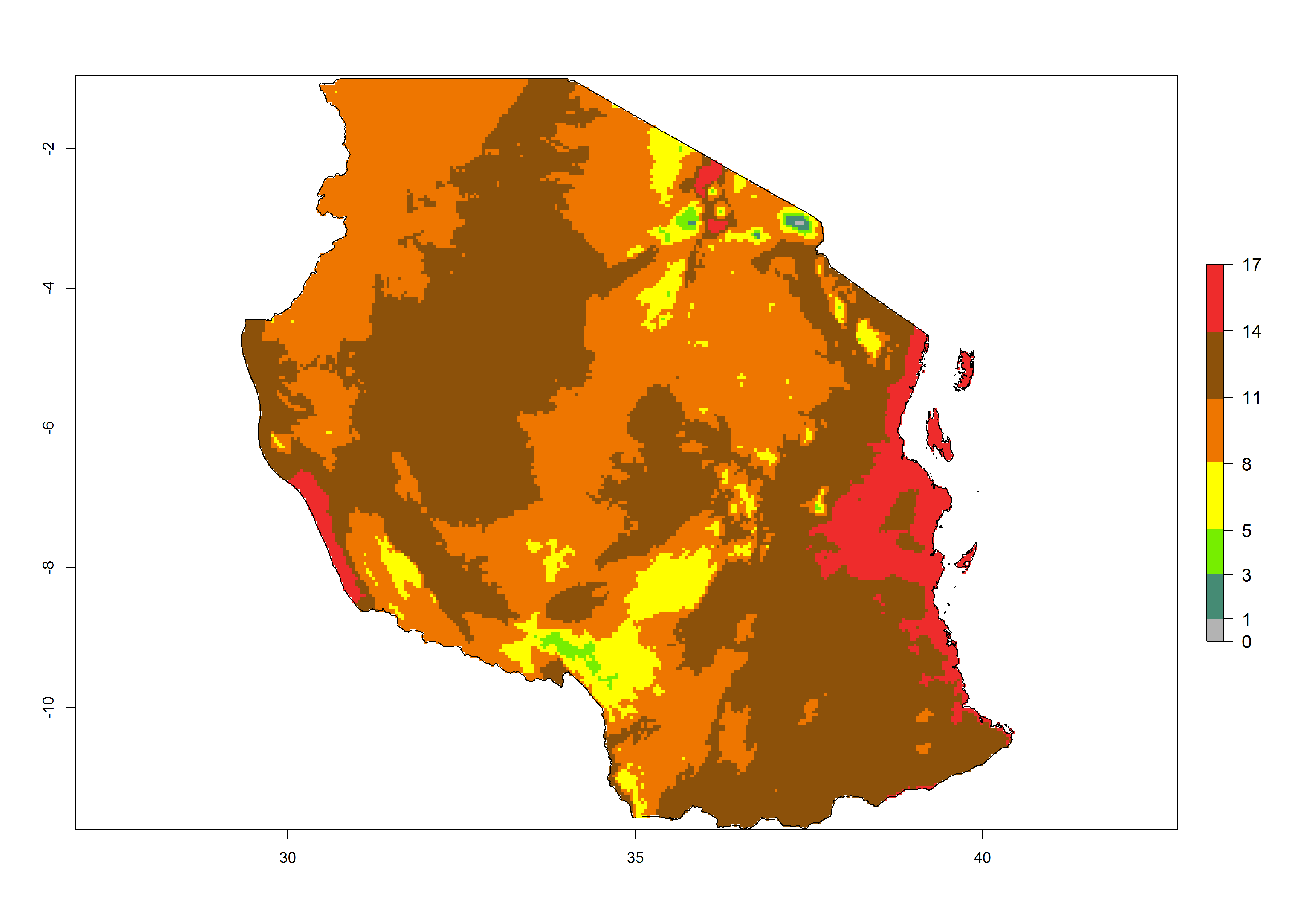 |
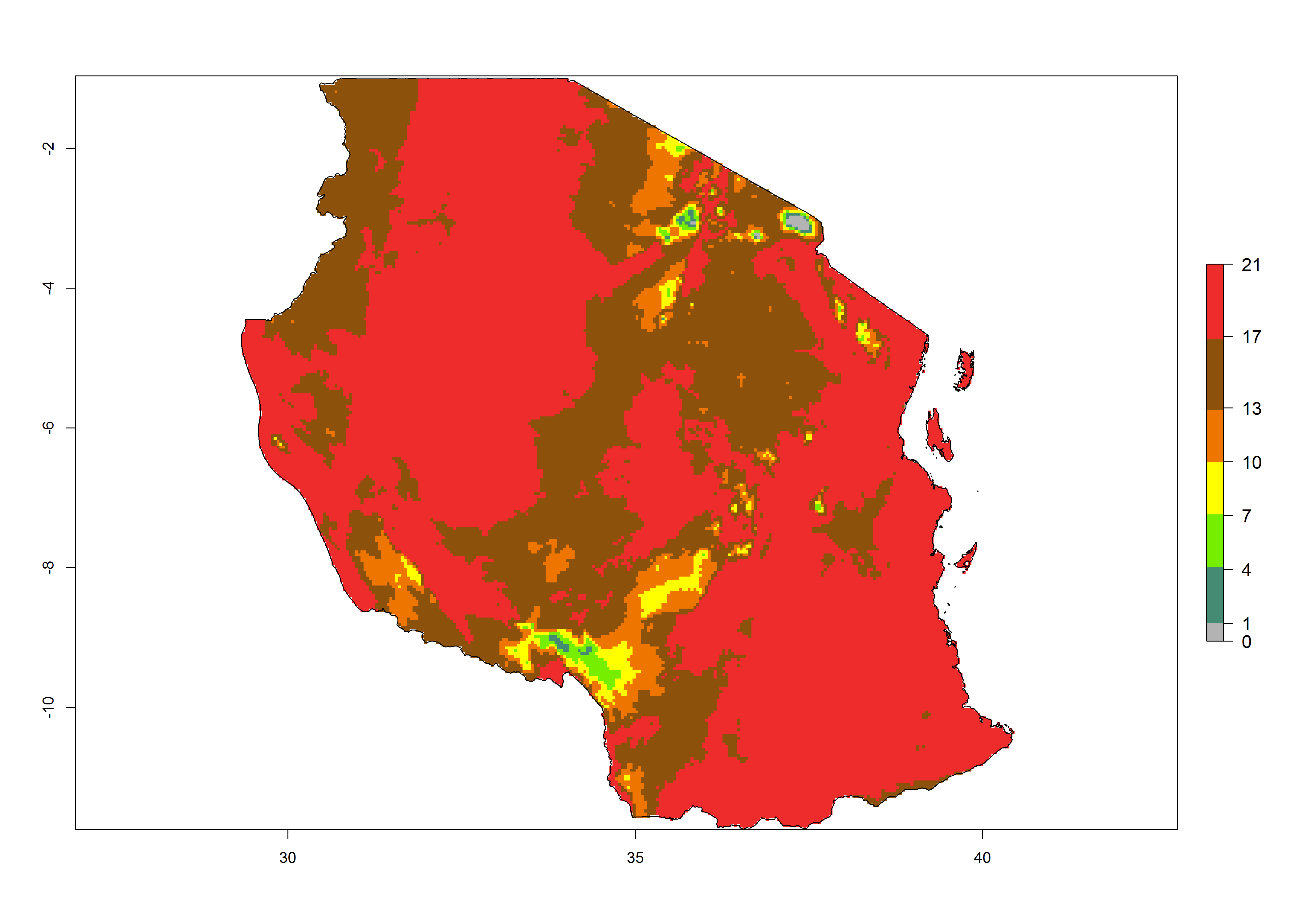 |
| k) Tunisia | ||
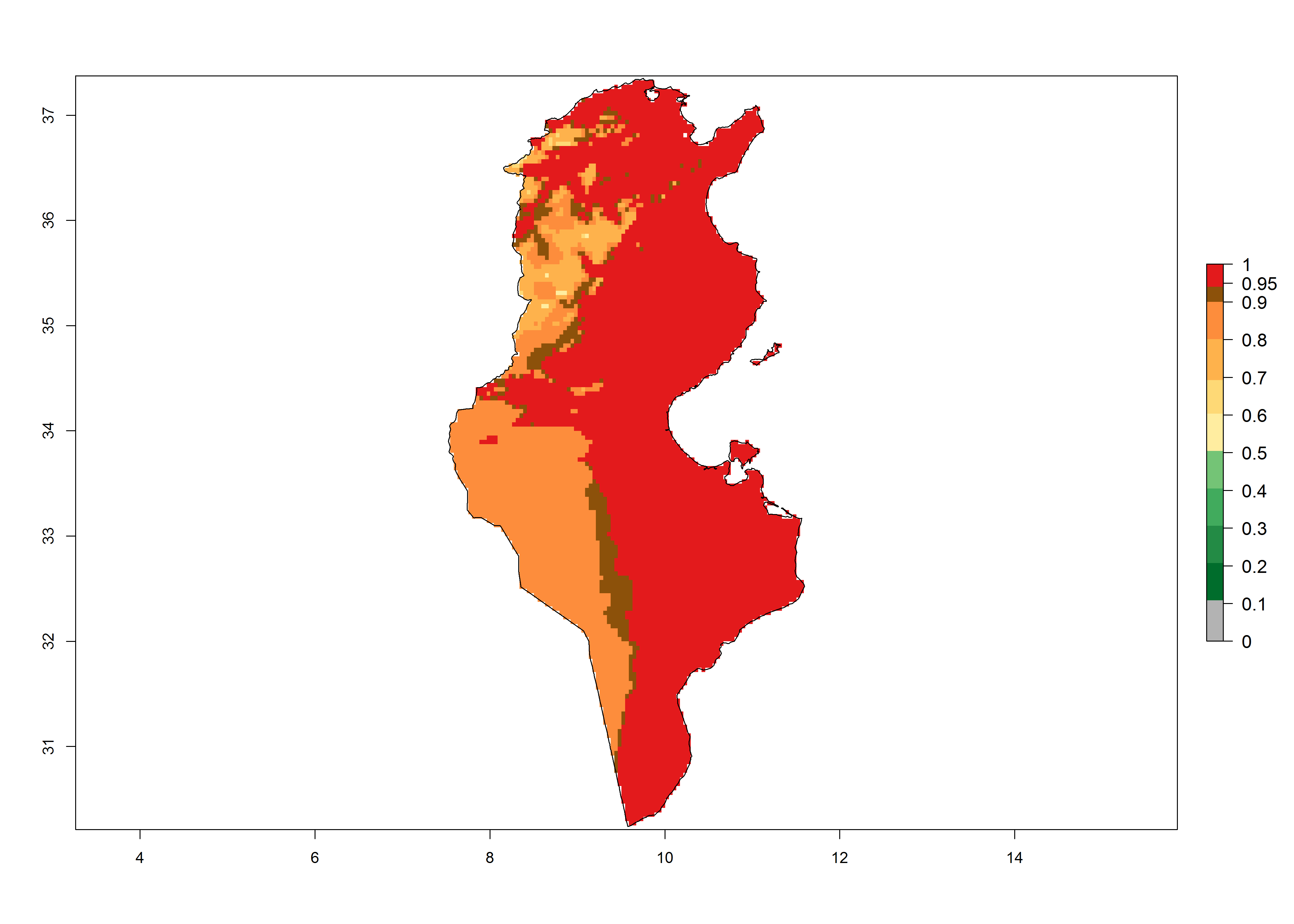 |
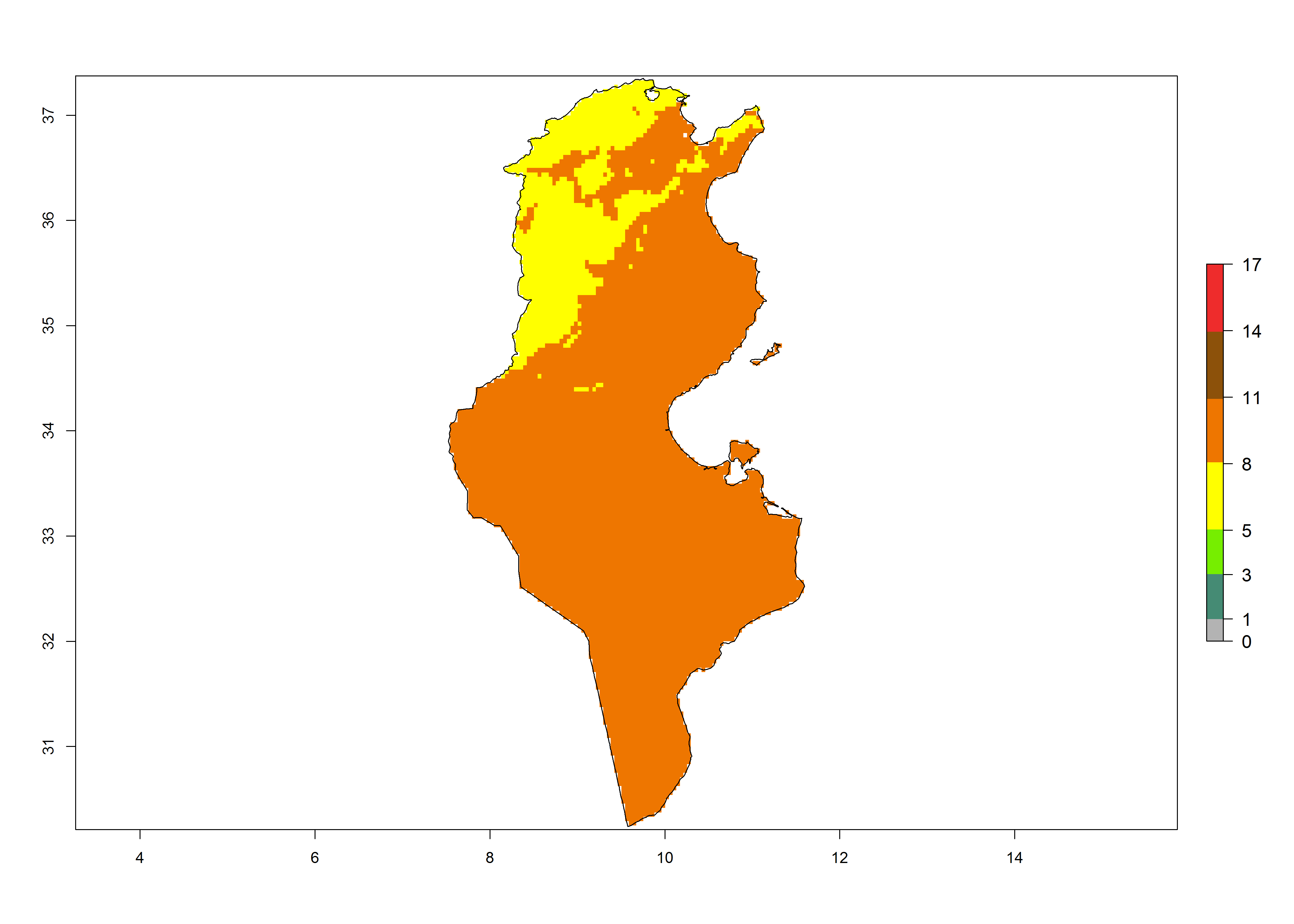 |
 |
| l) Uganda | ||
 |
 |
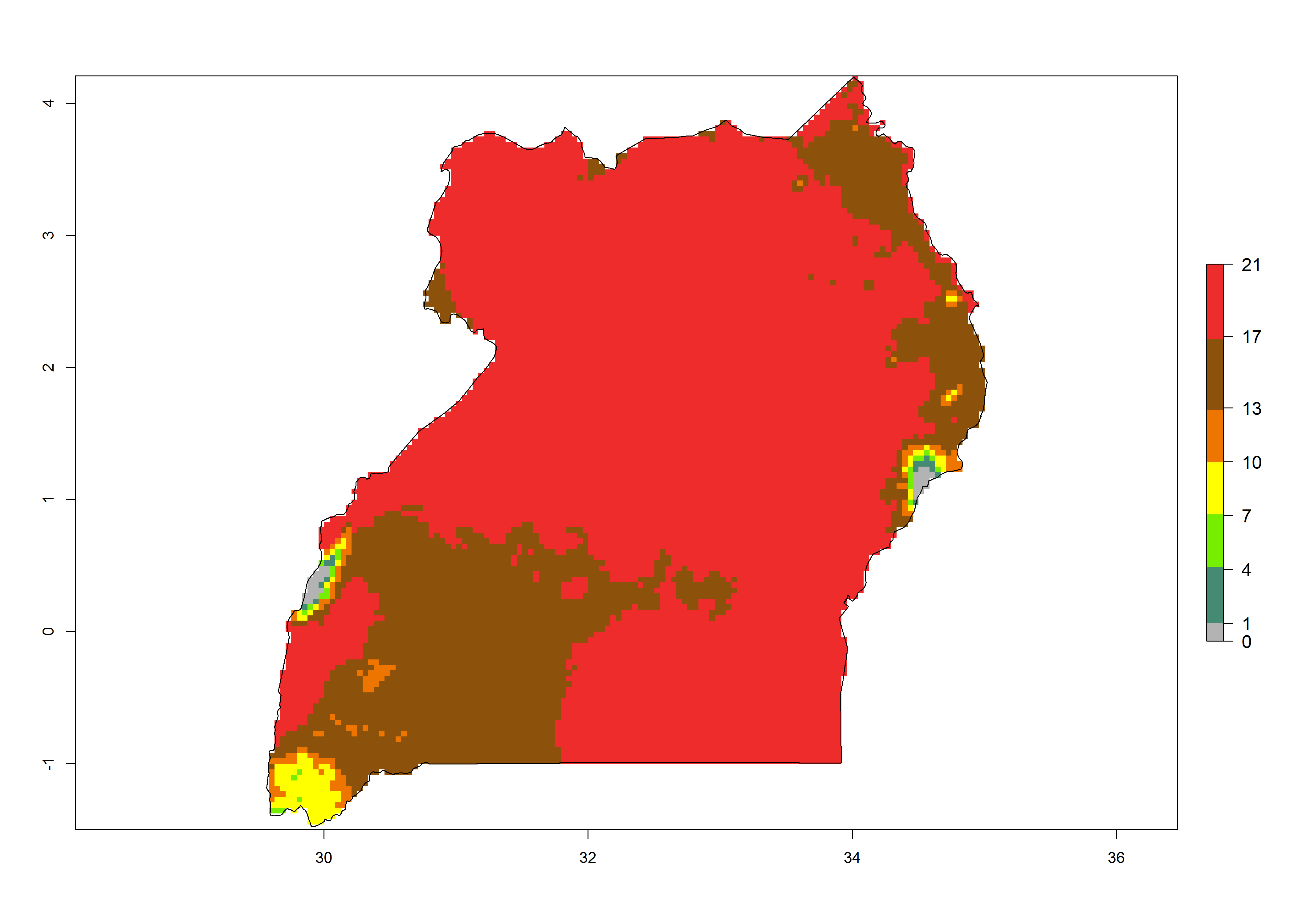 |
Figure 6. Establishment (ERI), abundance (GI, potential damage), and activity (AI, potential population growth) of the tomato leafminer, Tuta absoluta, in selected African countries according to model predictions for the year 2000. An ERI>0.7 is associated with potential permanent establishment.
Phytosanitary measures
- absoluta was added in 2004 to the EPPO A1 action list of pests recommended for regulation as quarantine pests. Plants for planting and fruits of tomato originating from countries where T. absoluta occurs should be free from the pest. Measures may include any combination of the following options: tomato and/or secondary host commodities must be imported only in commercial shipments, and must be produced in pest free facilities, for example, greenhouses or screen houses and packing houses approved by the NPPO of the country of origin. Internationally bound tomatoes must be packed in insect-proof boxes or in containers covered with an insect-proof mesh; greenhouses must have double doors and all ventilation systems and openings must be covered with a 1.6 mm mesh (or smaller) to prevent pest entry. Greenhouses must have traps with lures for Tuta absoluta at a minimum density of 2 traps per hectare or equivalent, with no less than two traps per greenhouse; and must be inspected by the NPPO during export season to look for T. absoluta. Each consignment of tomatoes and/or secondary host commodities must be accompanied by a phytosanitary certificate issued by the NPPO of the country of origin. The certificate must indicate that the commodity is free of soil, plant residues (no calyx, vines, stems or leaflet).
Adaptation to risk avoidance at farm level
IPM strategies developed include:
Monitoring with pheromone traps. Pheromone traps can be used both for early detection and monitoring the infestation density of the pest. It can also help to reduce the population when used for mass trapping, in which case a higher number of traps need to be placed for higher captures. For monitoring purposes, the number of traps placed should be 2-4 traps per hectare.
Biological control. Nesidiocoris tenuis (Miridae) is an efficient predator of eggs and small larvae of the tomato tuber moth. Egg parasitoids of the family Trichogrammatidae are the natural enemies most used worldwide in biological control programs against Lepidoptera, through inoculative and inundative releases. The application of T. pretiosum can reduce the population of T. absoluta, with a maximum percentage of control of 49% of the eggs. Therefore, it is suggested to combine with the application of a bio-insecticide based on Bacillus thuringiensis.
Biopesticides. The use of Neem oil (Azadiractin) acts as a contact and systemic insecticide against low infestations of Tuta absoluta larva when used on the tomato plant. The use of Bacillus thuringensis is recommended at low-medium infestation levels in combination with Azadiractin.
Cultural practices. Good agricultural practices and IPM should always be part of a growers’ practice for all types of pests and crops. Good agricultural practices for the control of T. absoluta include crop rotation with non-solanaceous crops (preferably cruciferous crops), ploughing, adequate irrigation and fertilization, removal of infested plants and complete removal of post-harvest plant debris and fruit. The removal of wild solanaceous host plants in the vicinity of the growing area is also encouraged, as these can host all stages of the pest which can then re-infect the growing crop.
Chemical control. Several insecticides have been registered for use against T. absoluta in potato; however this pest is known to develop resistance against effective insecticides. The Insecticide Resistance Action Committee (IRAC) suggests that alternating insecticides by their mode of action (manner in which an insecticide kills an insect: effect on nervous system, stomach, respiration, etc.) is more effective than alternating by active ingredient to delay the development of resistance.
Further reading
Barrientos ZR, Apablaza HJ, Norero SA & Estay PP (1998) [Threshold temperature and thermal constant for development of the South American tomato moth, Tuta absoluta (Lepidoptera, Gelechiidae).] Ciencia e Investigacion Agraria 25, 133–137 (in Spanish).
CABI. 2017k. Tuta absoluta datasheet. Invasive Species Compendium: Datasheets, maps, images, abstracts and full text on invasive species of the world. http://www.cabi.org/isc/datasheet/49260
CIP, 1996. Major potato diseases, ınsects and nematodes, 3rd edn. Centro Internacional de la Papa, Lima.
EPPO, 2017. EPPO Global database (available online). Paris, France: EPPO. https://gd.eppo.int/
EPPO. 2005. Tuta absoluta. Data sheets on quarantine pests. Bulletin OEPP/EPPO Bulletin 35, 434–435
Estay, P.2000. Polilla del tomate Tuta absoluta. Instituto de Investigaciones Agropecuarias (INIA-La Platina), http://www.inia.cl/medios/biblioteca/informativos/NR25648.pdf .
Garcia MF, Espul JC, 1982. Bioecology of the tomato moth (Scrobipalpula absoluta) in Mendoza, Argentine Republic. Revista de Investigaciones Agropecuarias, 17(2):135-146
Imenes SDL, Uchôa-Fernandes MA, Campos TB, Takematsu AP, 1990. Aspectos biológicos e comportamentais da trata do tomateiro Scrobipalpula absoluta (Meyrick, 1917), (Lepidoptera-Gelechiidae). Arquivos do Instituto Biológico, Sao Paulo, 57(1/2):63-68.
Niederwieser F. 2017. Guidelines for the control of tomato leafminer (Tuta absoluta) on potato.
Notz, A.P., 1992. Distribution of eggs and larvae of Scrobipalpula absoluta in potato plants. Rev. Facul. Agron. (Maracay), 18: 425-432.
OIRSA. 2015. Manual de procedimientos para la vigilancia, prevención y control de la polilla del tomate Tuta absoluta (Lepidóptera: Gelechiidae) en la región del OIRSA
SENASICA. 2016. Palomilla del tomate Tuta absoluta Meyrick. Ficha Técnica No. 28, 71 pp.
Vargas HC, 1970. Observaciones sobre la biologfa y enemigos naturales de la polilla del tomate, Gnorimoschema absoluta (Meyrick) (Lepidoptera: Gelechiidae). Idesia (Chile), 1:75-110
Tropea Garzia, G., Siscaro, G., Biondi, A. and Zappalà, L. (2012), Tuta absoluta, a South American pest of tomato now in the EPPO region: biology, distribution and damage. EPPO Bull, 42: 205–210. doi:10.1111/epp.2556