4.1.2 Potato Pests / Guatemalan potato tuber moth, Tecia solanivora (Povolny 1973)![]()
Synonym: Scrobipalpopsis solvanivora (Povolny 1973)
Taxonomic position: Insecta, Lepidoptera, Gelechiidae
Authors: B. Schaub, P. Carhuapoma, & J. Kroschel
Common names
Guatemalan potato tuber moth (English), polilla guatemalteca de la papa, polilla gigante de la papa (Spanish)
Hosts
Potato (Solanum tuberosum L.)
Detection and identification
Larvae of Tecia solanivora feed exclusively on tubers during potato cultivation and, after harvest, in potato stores. Damage is caused by larvae that bore galleries into the tubers, making them unsuitable for consumption (Photo 1A). In potato fields T. solanivora attack occurs from tuberization until harvest. Eggs are laid on the soil close to the potato plants; probably due to their sticky surface they get camouflaged by soil particles and cannot be seen. Tuber infestation caused by first instar larvae can be hard to detect, such that even with precautionary measures infested tubers are transferred to potato stores, where the pest and infestation of the whole stock can propagate further. As larvae develop inside the tubers, feces become visible around the entry hole and indicate infestation. Inside the tubers larvae bore irregular galleries that may run into the interior of the tubers or remain directly under the skin. After the larvae have left the tubers, the exit hole is clearly visible (Photo 1B).
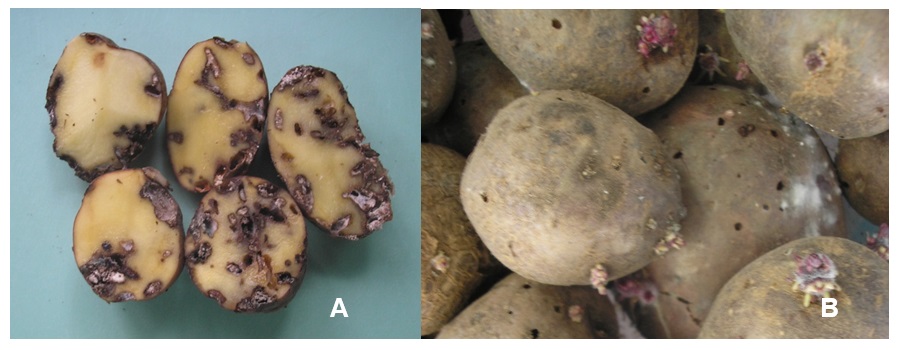
The mining causes tubers to lose weight, which is exacerbated by increased transpiration through the wounds, causing them to shrink. The wounds also provide entry points for microorganisms and can cause secondary infestation with pathogens. Infestation of stored tubers can result in total destruction; however, this largely depends on the length of the storage period and the prevalent storage temperature that determines how fast T. solanivora can reproduce and build up several generations.
Morphology
Egg
Eggs are laid in groups (Photo 2A) and are oval shaped and 0.5 x 0.4 mm in size. The color of the eggs turns from pale white to yellow within a few days. Before hatching, eggs appear black as the dark head capsule of the larva becomes visible.
Larva
First instar larvae are 2–3 mm long; the body is a pale, transparent white and the head capsule and pronotum are dark brown (Photo 2A). Second instar larvae are no longer transparent and have some small, dark brown spots along their entire body. Third instar larvae look similar but are larger in size; fourth instar larvae are purple on their dorsal part and are green on their ventral part (Photo 2B). Three pairs of thoracic podia and five pairs of pseudopodia are now clearly visible. Larvae reach a length of 12–17 mm and a diameter of 2–3 mm. Last instar larvae leave the potato tubers in search of a protected pupation site. Before pupation, feeding is stopped, movements are reduced, and larvae shrink in size.
Pupa
The pupae are 7–10 mm long; female pupae are bigger than males (Photo 2C). Their color first turns from light brown to dark brown just before emergence. In many cases, they can be found within a cocoon, which, depending on the environment, might include soil particles.
Adult
Adults reach a size of 12–18 mm; females (Photo 2D) are bigger than males (Photo 2E). Their color is brownish to gray; males are darker than females. The wings are folded to form a roof-like shape. The pattern on the forewings is characterized by a dark line from the front to the back that is interrupted by two dark spots surrounded by a bright circle.

Biology
Adult females lay their eggs on the soil close to the potato plant; few eggs are laid on the potato plant itself. The hatching larvae pass through the soil and enter the potato tubers. They feed and bore galleries throughout the tubers until they reach the fourth instar larva stage. Mature larvae leave potato tubers and pupate on the soil surface. When infested potato tubers are stored, the life cycle continues inside of potato stores. The larvae pupate between potato tubers, on potato bags, or in other niches inside the storage facilities. The emerging adults mate and the females lay eggs on the surface of the potato tubers.
Temperature-dependent development
Development of all T. solanivora life stages is possible at 9°–28°C (see Annex 7.3.2). At 10°C, the median immature development time was 208 days. It decreased with rising temperatures and was shortest with 37 days at a temperature of 27°C. Lifespan of adults decreased from about 40 days at 10°C to about 10 days at temperatures above 25°C. At temperatures below 20°C, males lived on average 37% longer than females. Mortality of eggs was around 10% at temperatures of 10°–25°C, increased sharply at higher and lower temperatures, and reached 100% at 5°–30°C. For larvae, mortality was lowest at 15°–20°C; pupae mortality was lowest at 17°–21°C. Oviposition peaked at 15°C with 311 eggs per female; at temperatures above 25°C, fewer than 50 eggs were laid, and at 4.5°C no eggs were laid. At all temperatures most females survived until 75% of the eggs were laid.
The functions that had been established to describe the development time and rate, mortality, and reproduction were compiled into an overall phenology model. The following life-table parameters were calculated: the intrinsic rate of increase (rm) and the finite rate of increase (λ) indicate a population growth between 9°C and 28°C, with peaks between 19°C and 23°C, where a daily population growth of 7.0% was estimated. At this temperature range, doubling time (Dt) was shortest with 10 days. The mean generation time (T) decreased with temperature and was shortest at 27°C, with 37 days from egg to egg. The gross reproduction rate was highest at 14°–15°C, with 197 female offspring/female. When mortality of females before reproduction was considered, the net reproduction rate (Ro) was highest at 15°–17°C, with around 65 female offspring/female.
Means of movement and dispersal
The spread of T. solanivora from Central America southwards was linked in several cases to importation of infested seed. In 1970 Costa Rica imported infested seed from Guatemala, and in 1983 Venezuela was invaded by the pest through seed tubers imported from Costa Rica. When T. solanivora reached Ecuador, it spread within 2–3 years to almost all potato-growing zones in the country, probably facilitated by the transportation of infested seed tubers. Dispersal by flight of adults probably does not occur over large distances, since female moths tend to sit on the ground and show low flight activity throughout the day.
Economic impact
The damage observed at harvest varies with the climatic conditions: low temperature and rainfall reduce T. solanivora reproduction and survival. During dry seasons tuber damage rates of 2–15% are observed in the Andean potato production. Complete harvest losses have been observed occasionally, mainly after the invasion of new areas when farmers were not yet familiar with pest control or when fields were abandoned due to overproduction and low market prices. When infested potato tubers are stored without application of control methods, T. solanivora can destroy the whole potato stock, depending on the storage period and temperature.
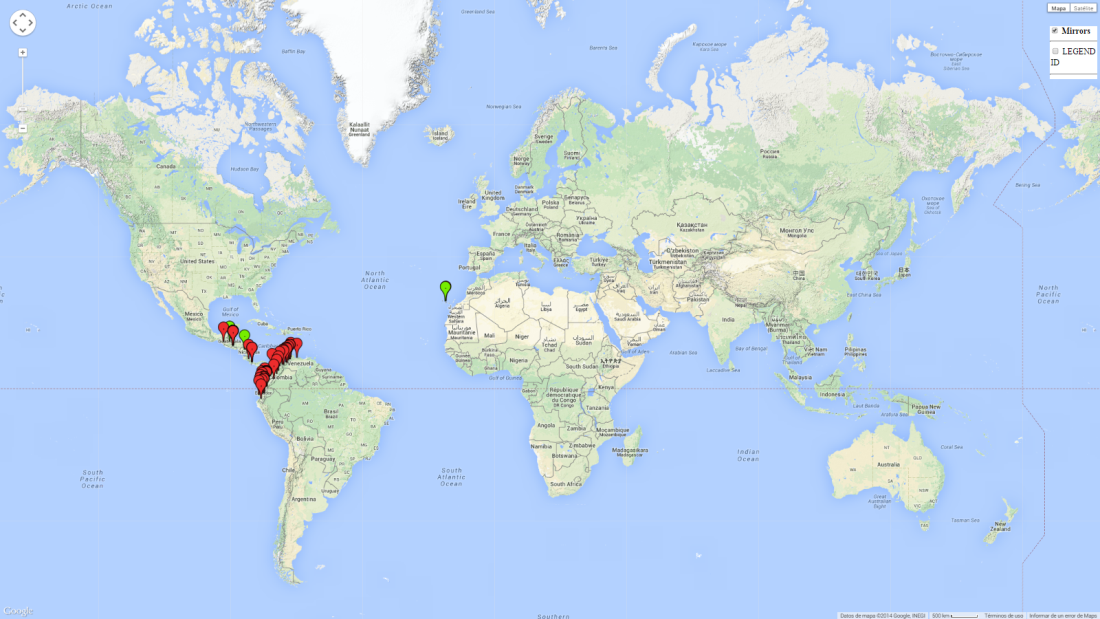
Geographical distribution
T. solanivora originated in Guatemala, and its distribution was limited to this part of Central America until seed trade introduced the pest into Costa Rica in 1970 and into Venezuela in 1983. From there it spread to Colombia and Ecuador. In 1999, T. solanivora was detected in Tenerife, Canary Islands, and in 2010 it was reported for the first time in southern Mexico (Fig. 1).
| Europe | Canary Islands (Spain) |
| Central America | Costa Rica, El Salvador, Guatemala, Honduras, Mexico, Nicaragua, Panama |
| South America | Colombia, Ecuador, Venezuela |
Phytosanitary risks
T. solanivora is currently present in potato-growing zones with subtropical climate in Central America, northeastern parts of South America, and the Canary Islands. Since the distribution of T. solanivora is still limited to certain regions, the prevention of distribution to further areas at risk is essential. As potato is the only host crop of T. solanivora, its further distribution as in previous cases will very likely also be strongly linked to trade and the formal and informal movement of seed tubers. Hence, neighboring countries such as Peru, where potato is a major crop, are especially at high risk. As happened on the Canary Islands, distant countries might also be invaded when infested potato tubers are traded. The European Plant Protection Organisation (EPPO) included T. solanivora in its A2 List of pests recommended for regulation as quarantine pests, as T. solanivora is already present on the Canary Islands. The North American Plant Protection Organization lists T. solanivora in its Phytosanitary Alert System.
Risks mapping under current and future climates
Global Risks
Changes in establishment and future distribution
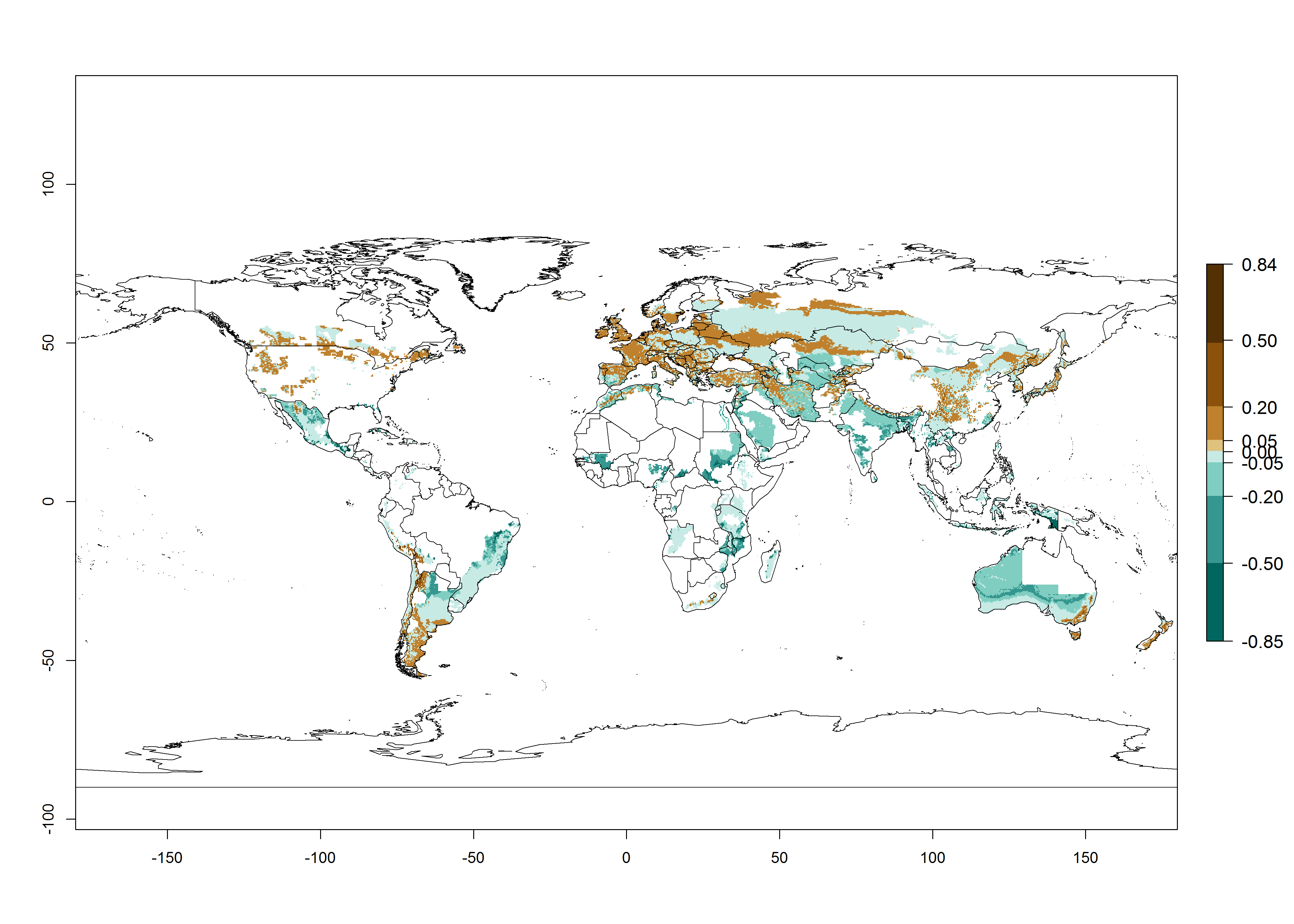
Potato-growing areas of Central and South America infested with T. solanivora have an establishment risk index (ERI)>0.95. Therefore, regions with an ERI>0.95 are considered to be at risk of T. solanivora establishment. In the Americas, establishment is potentially possible in parts of the potato-growing zones from about 30° north to 40° south; in Africa, the potato cultivation regions in sub-Saharan countries and at the Mediterranean coast are at risk. In Asia, T. solanivora might establish in all potato-growing zones from 10° north to 10° south and in the higher elevated growing zones up to 30° north. In Australia, temperature conditions in the southern part would allow establishment. These geographies at risk are basically the same in the years 2000 and 2050, but the total area at risk decreases slightly (Fig. 2). An increase in the risk is estimated for temperate regions beyond 35° north or 40° south and in high-elevated tropical growing zones of South America. In the lower potato-growing zones in the tropics and subtropics, the risk of establishment might potentially decrease. In general, T. solanivora is currently thought to spread only in potato-growing zones where temperature conditions allow it to develop continuously throughout the year. There is no experience on the potential establishment in potato-growing regions with seasonal climate. Furthermore, in the current distribution zone of T. solanivora, potatoes are cultivated at high altitudes in the Andes, and cold temperatures limit its further distribution into even higher cultivated zones. At lower zones, where warm temperatures might limit T. solanivora population growth, potatoes are not cultivated in the Andes. From the warmer Central American potato-growing zones, no upper temperature limits for T. solanivora establishment are reported. Finally, rainfall patterns and crop management are further important factors that impact pest establishment.
 |
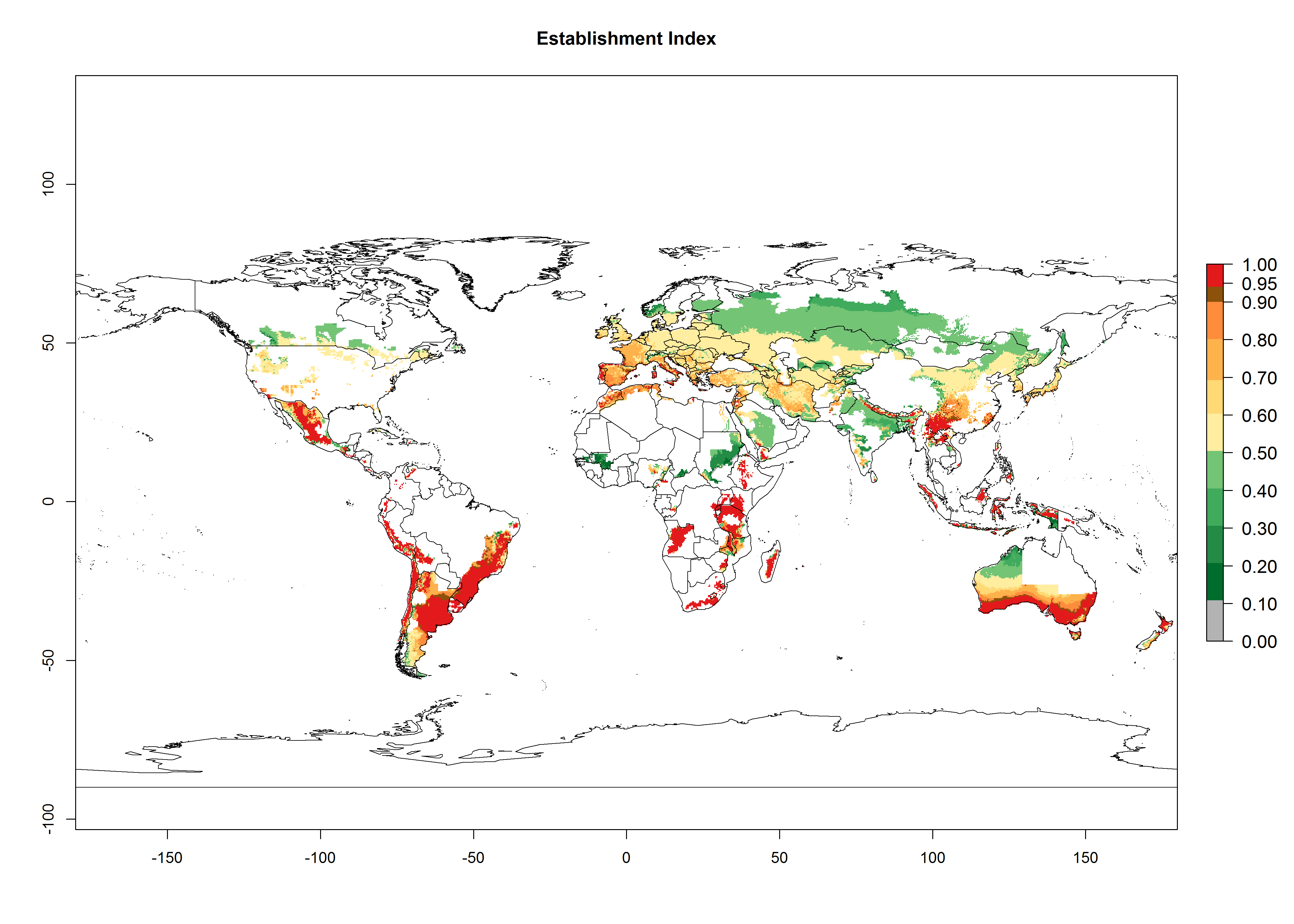 |
Figure 2. Changes in establishment and potential distribution of the Guatemalan potato tuber moth, Tecia solanivora, in potato production regions worldwide according to model predictions, using the ERI for the years 2000 (A) and 2050 (B), and changes of the ERI between 2000 and 2050 (C). An ERI>0.95 is associated with potential permanent establishment.
The number of generations predicted for the potato-growing areas with existing T. solanivora infestation range mainly from 3 to 6; in some parts of Central America up to 8 generations might develop. There are few reports on the number of generations that develop at specific geographies for validation of the simulations. However, the longest development time observed under natural conditions is about 4 months; lower temperatures would result in high immature mortality. The shortest development times reported under fluctuating temperature conditions in the Andes are about 2 months. Therefore, the predictions of 3–6 generations for most of the infested potato-growing zones, where climatic conditions do not change much throughout the year, are in accordance with observed generation cycles. For these growing zones, an activity index (AI) of 2–6 was estimated. Especially in Central America there are areas with an AI of 10–11.5; however, in these warmer regions potato cultivation is less important and might be limited to the cooler season of the year.
Changes in abundance
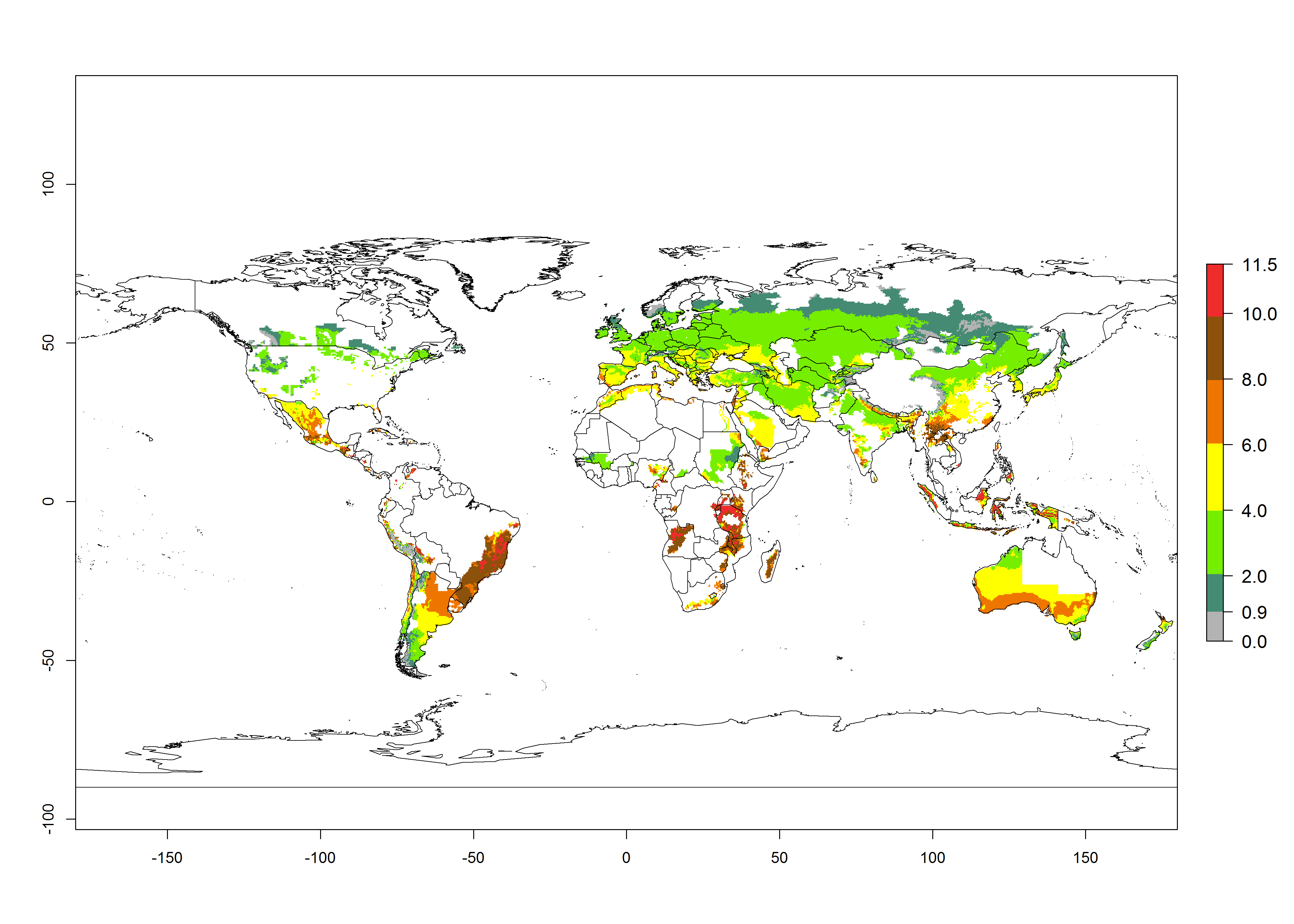
In the African and Asian potato production regions, where establishment of T. solanivora might be possible, frequently a generation index (GI)>6–10.5 generations per year has been estimated (Fig. 3). Also, the activity would be higher in many parts of these regions with an AI>6–11.5. For the year 2050 a further increase by 1–2 generations per year is predicted; the AI, however, would potentially decrease in the tropical zone. In the temperate regions and in the high-elevated tropical potato-growing regions, the AI is predicted to increase from the years 2000 to 2050 by values up to 3.
| GI | AI | |
| 2000 | 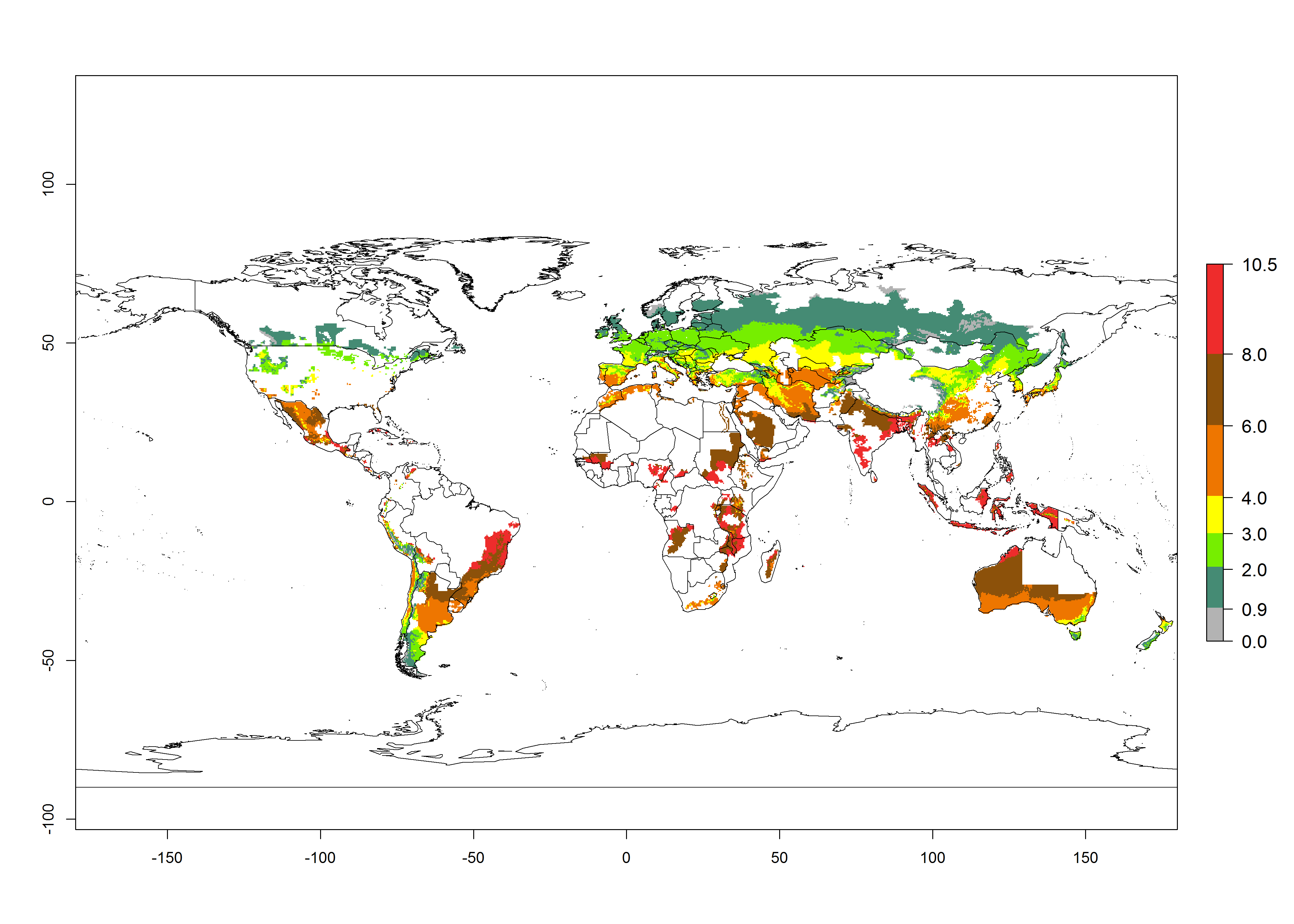 |
 |
| 2050 | 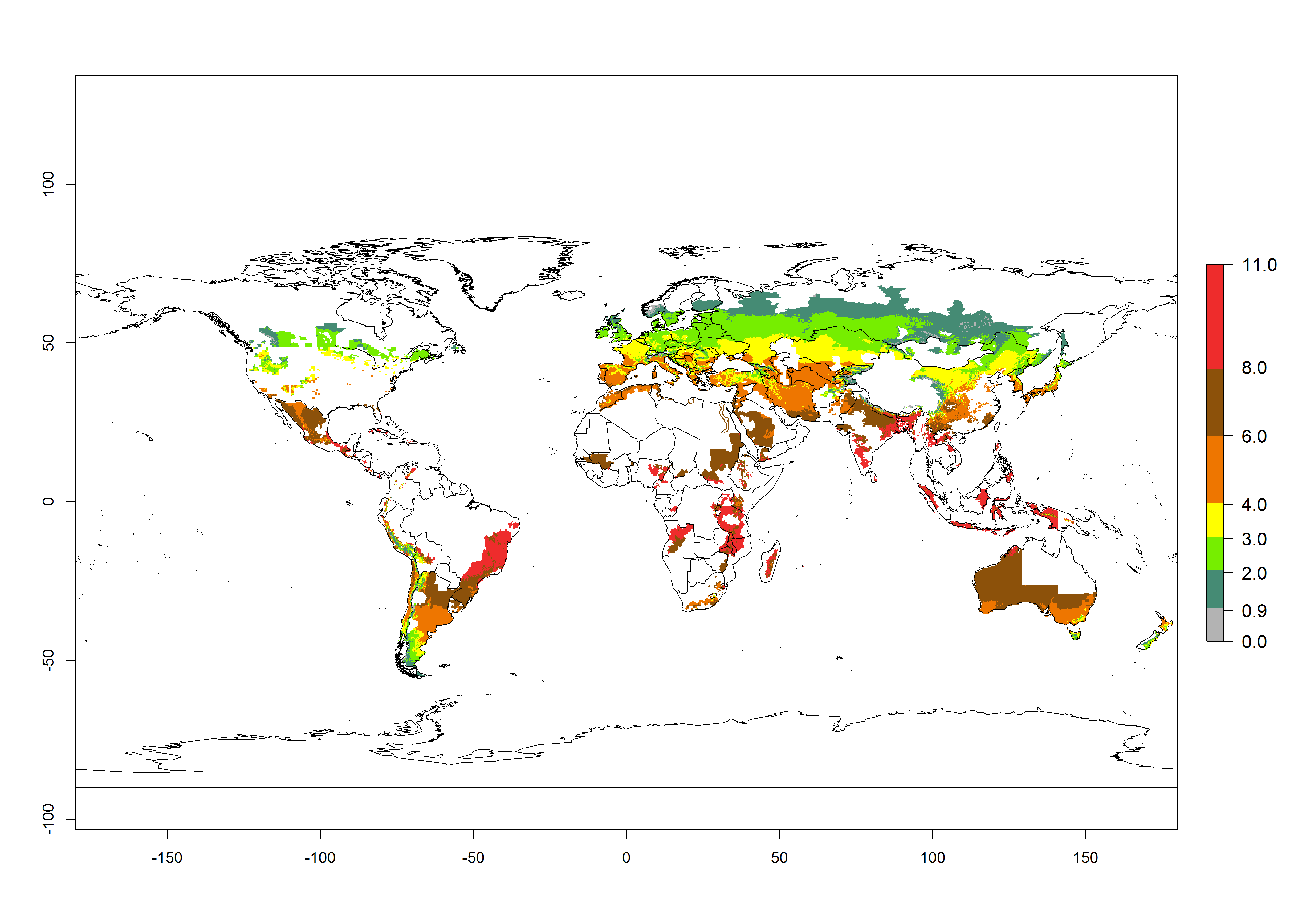 |
 |
| Index change (2000 – 2050) | 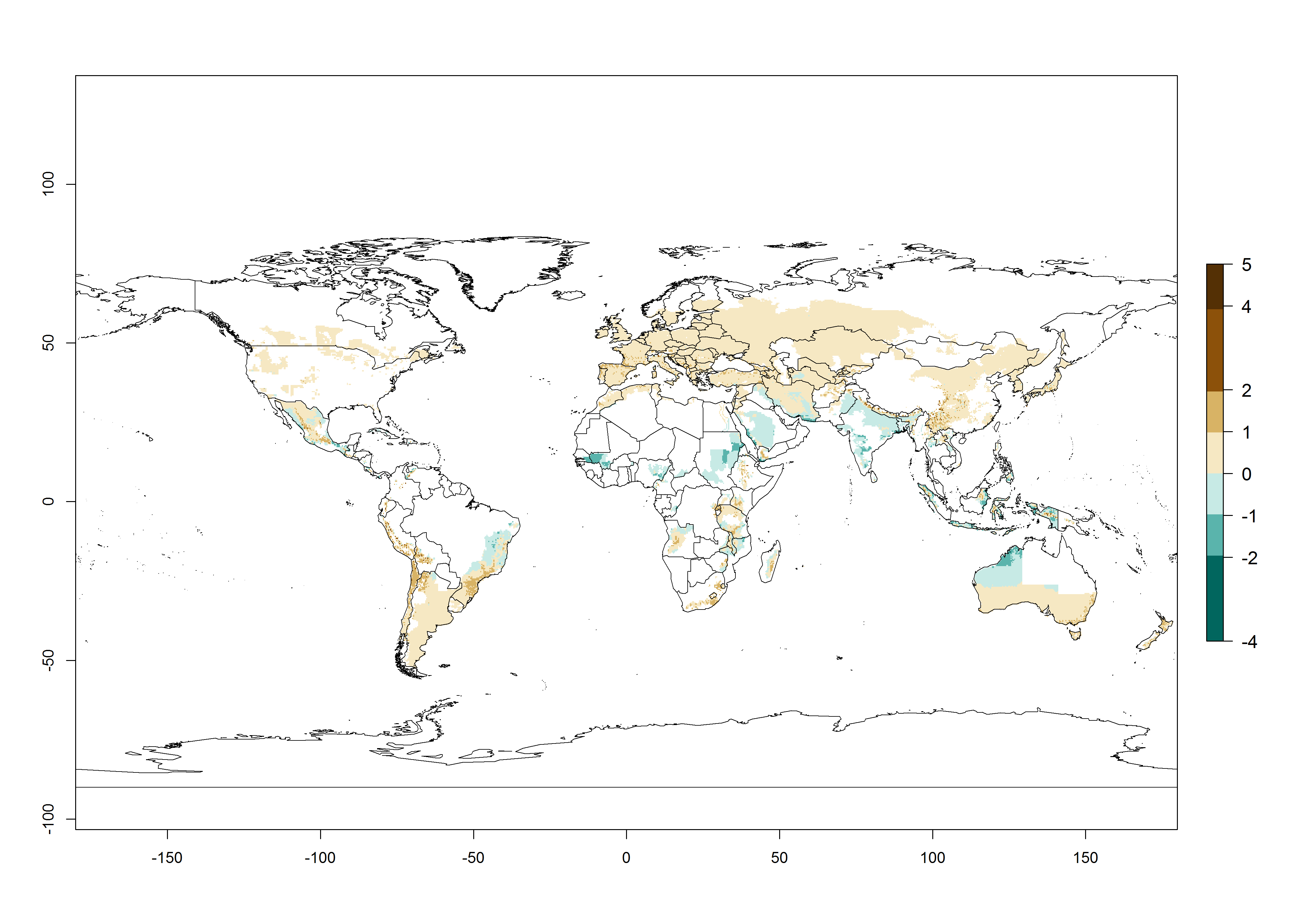 |
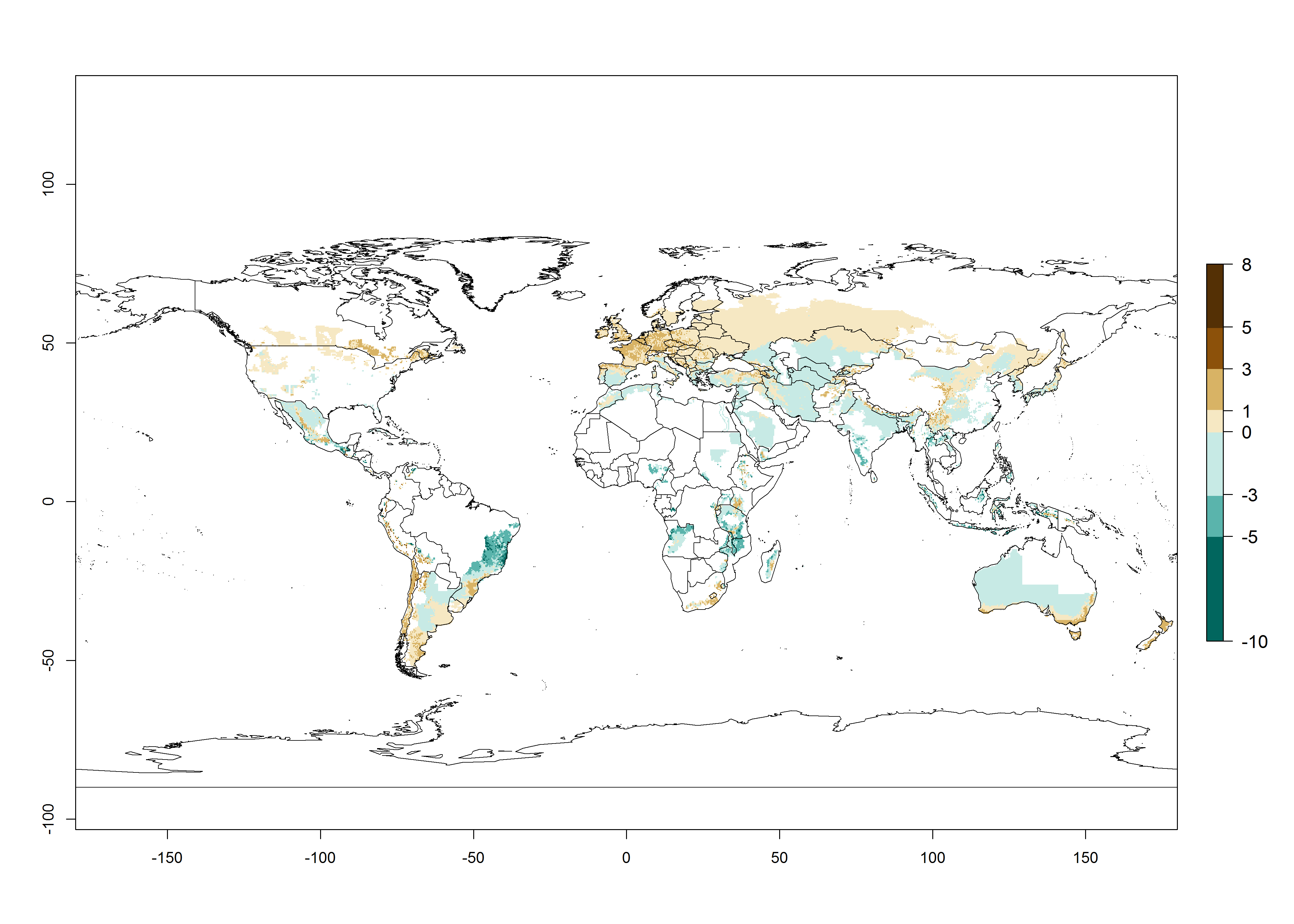 |
Figure 3. Changes in abundance (GI, damage potential) and activity (AI, potential population growth) of the Guatemalan potato tuber moth, Tecia solanivora, in potato production systems worldwide according to model predictions, using the GI (A, B) and the AI (D, E) for the years 2000 and 2050, and the absolute index change (C, F).
Regional Risks for Africa
Changes in establishment and future distribution
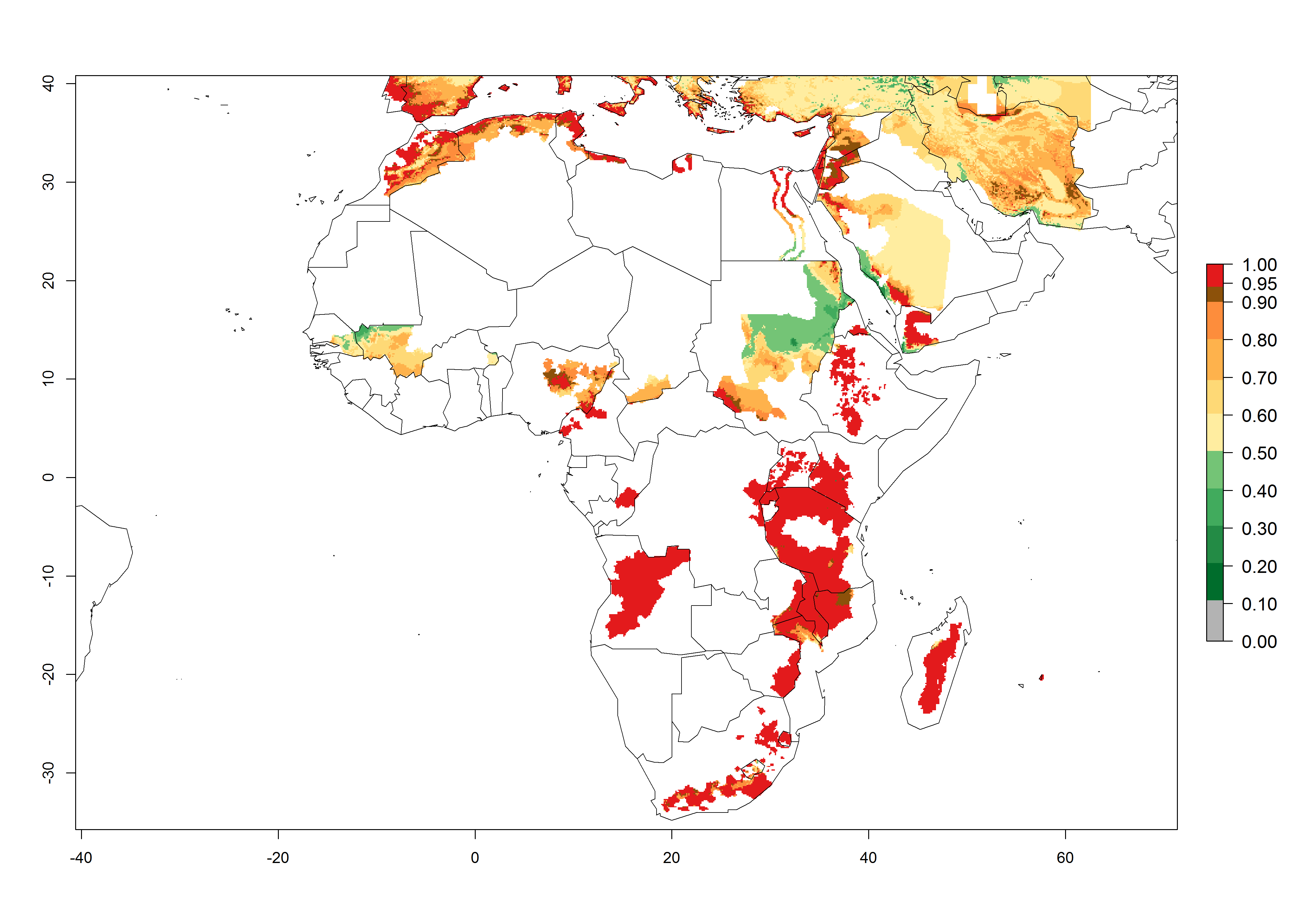
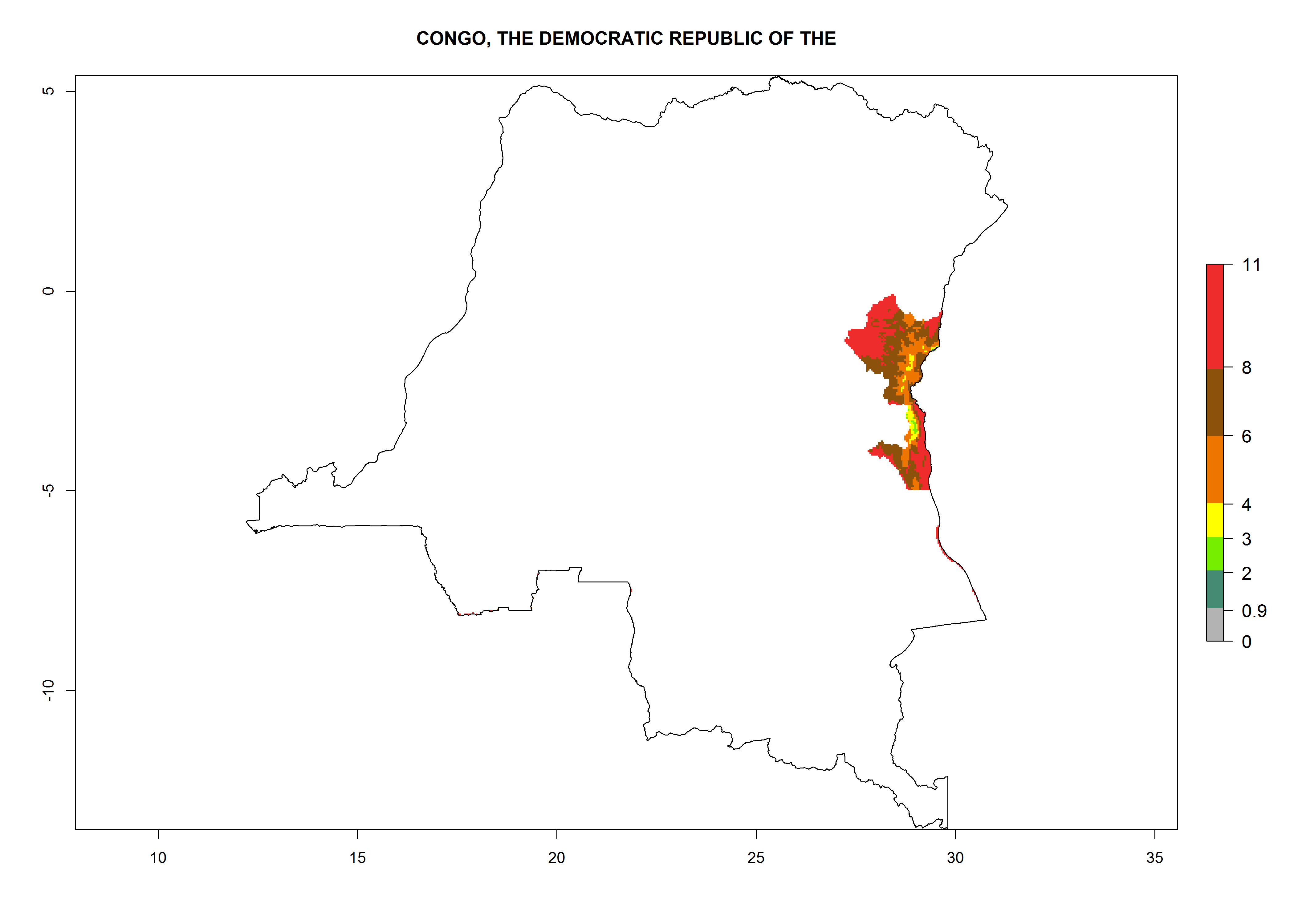
The African countries, where temperature conditions might allow establishment of T. solanivora, are far away from the infested regions in Central and South America and the Canary Islands. T. solanivora might reach these regions only by accidental introduction of infested potato tubers. According to FAOSTAT 2014, potato tubers were not imported from infested countries to the African countries with an ERI>0.95 during the past 10 years. For the year 2000, an ERI>0.95, which would allow establishment, is estimated for considerable parts of potato-growing regions of countries at the Mediterranean coast of North Africa (Morocco, Algeria, Tunisia) and in Ethiopia, Kenya, Uganda, Tanzania, Rwanda, Malawi, Angola, Congo, DR Congo, Cameroon, Zimbabwe, Zambia, Mozambique, South Africa, Swaziland, and Madagascar (Fig. 4).

 |
 |
Figure 4. Changes in establishment and potential distribution of the Guatemalan potato tuber moth, Tecia solanivora, in African potato production systems according to model predictions, using the ERI for the years 2000 (A) and 2050 (B), and changes of the ERI between 2000 and 2050 (C). An ERI>0.95 is associated with potential permanent establishment.
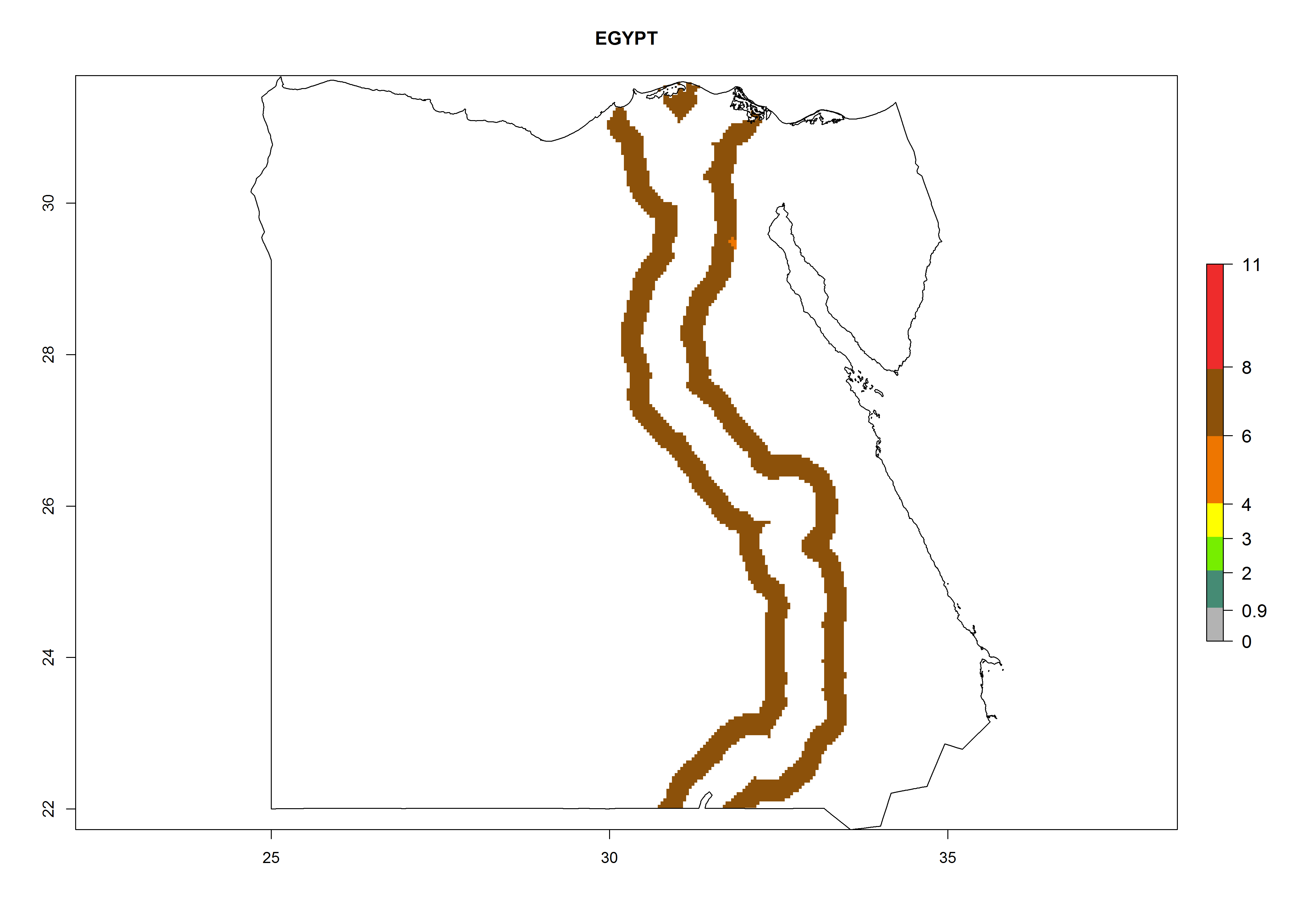
For 2050, the risk potentially increases in Morocco and South Africa in some higher elevated potato-growing zones. For Egypt, Tunisia, Algeria, Congo, Zambia, and Mozambique, the risk of establishment will potentially reduce.
Changes in abundance
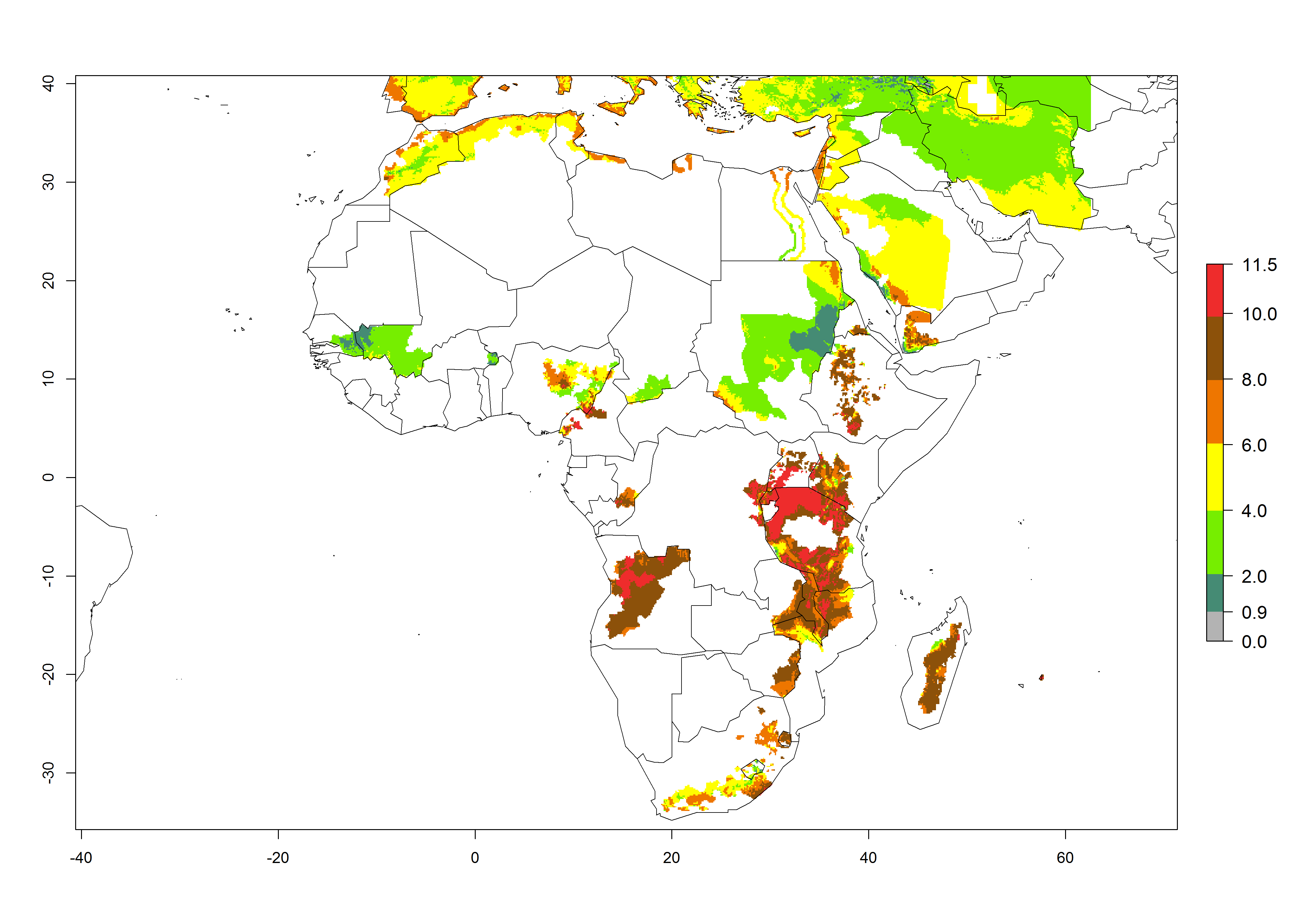
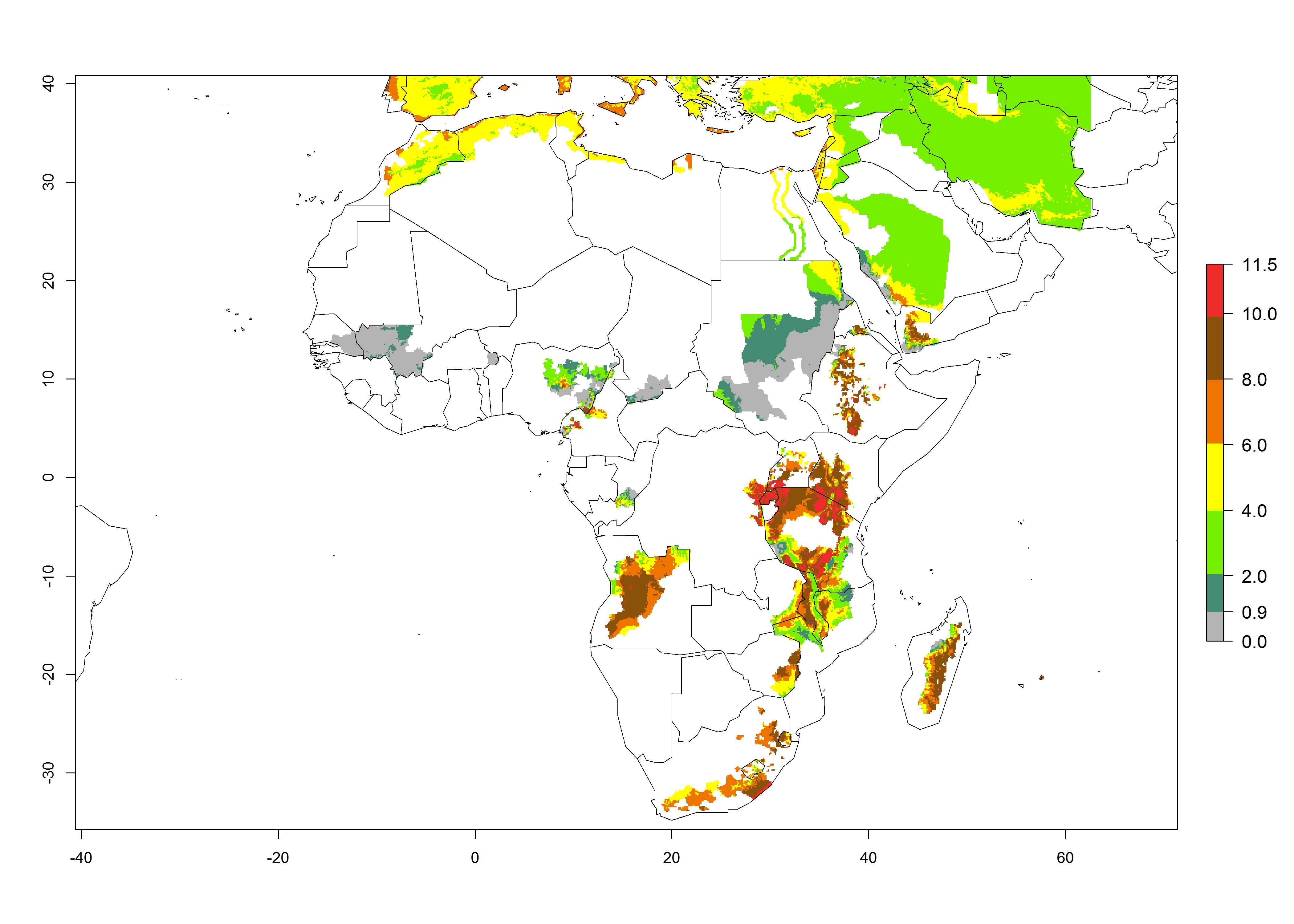
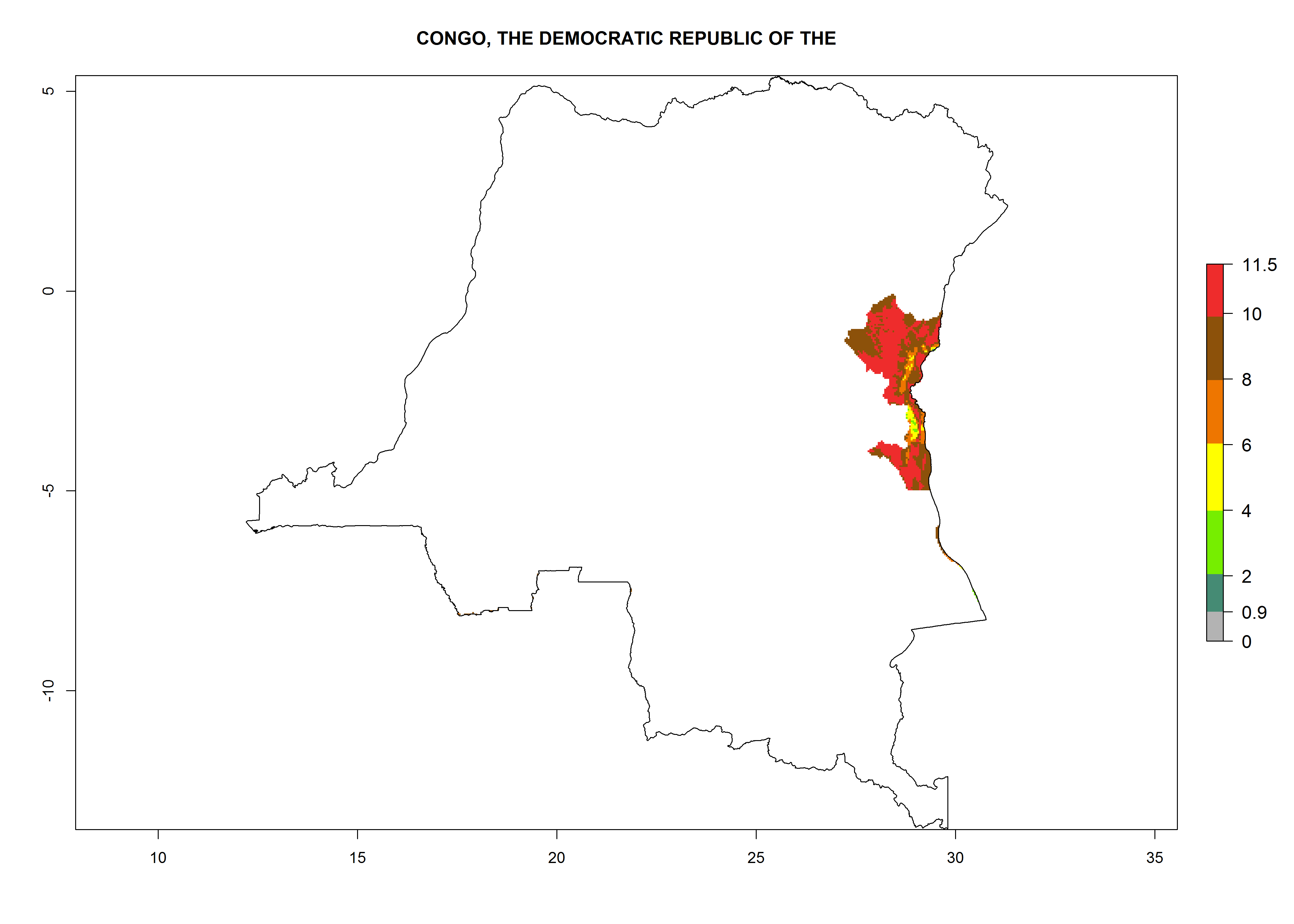
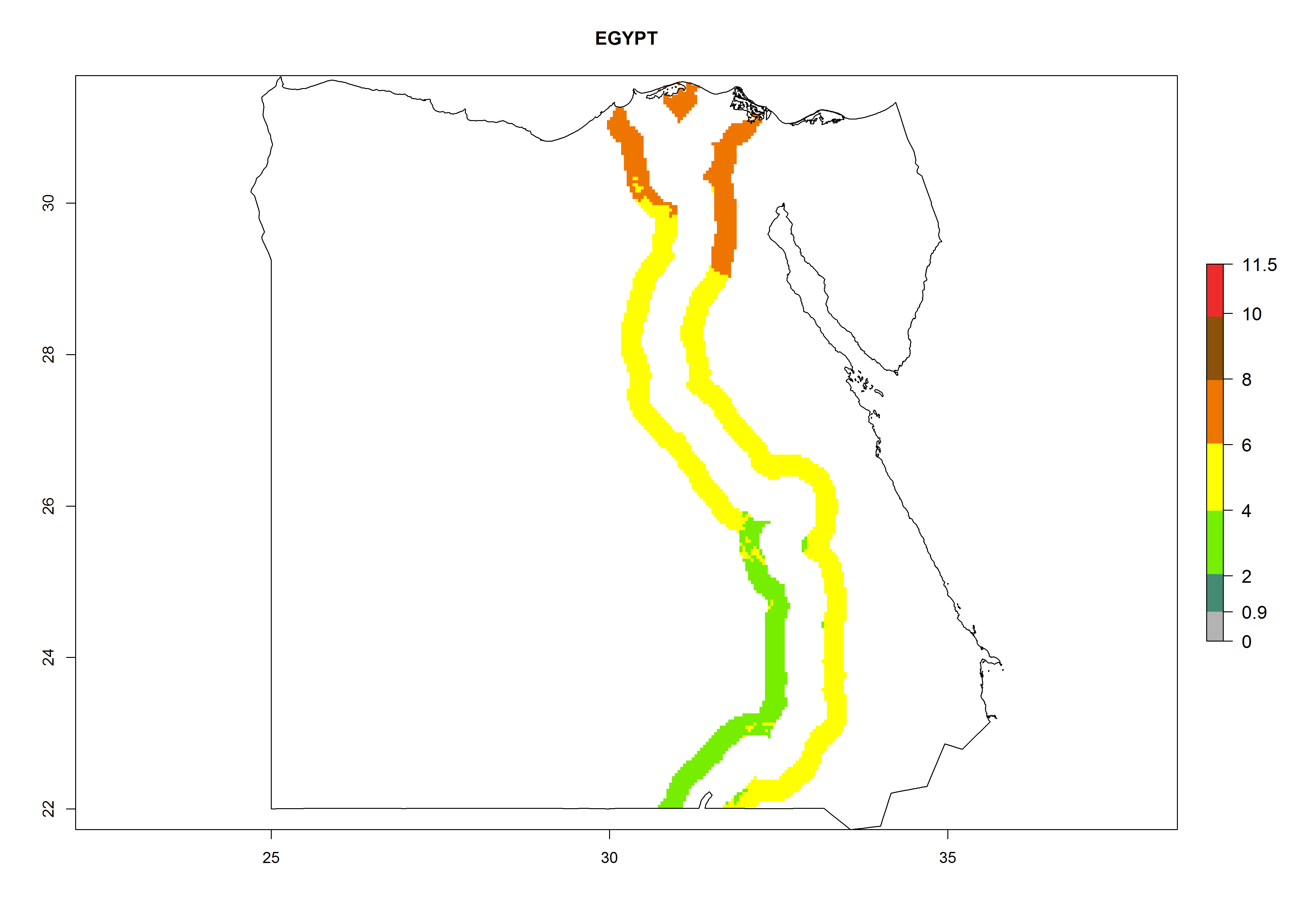
A GI>6–10.5 is predicted for most of the areas at risk; in most of South Africa 4–6 generations are estimated per year. Until 2050 the number of generations still increases, and the highest possible number of 8–10.5 generations per year is reached in large parts of the potato-growing zones. However, at these temperatures, mortality is high and the AI decreases by 2–4 values in most parts except for South Africa and Morocco, where the risk increases (Fig. 5).
| GI | AI | |
| 2000 | 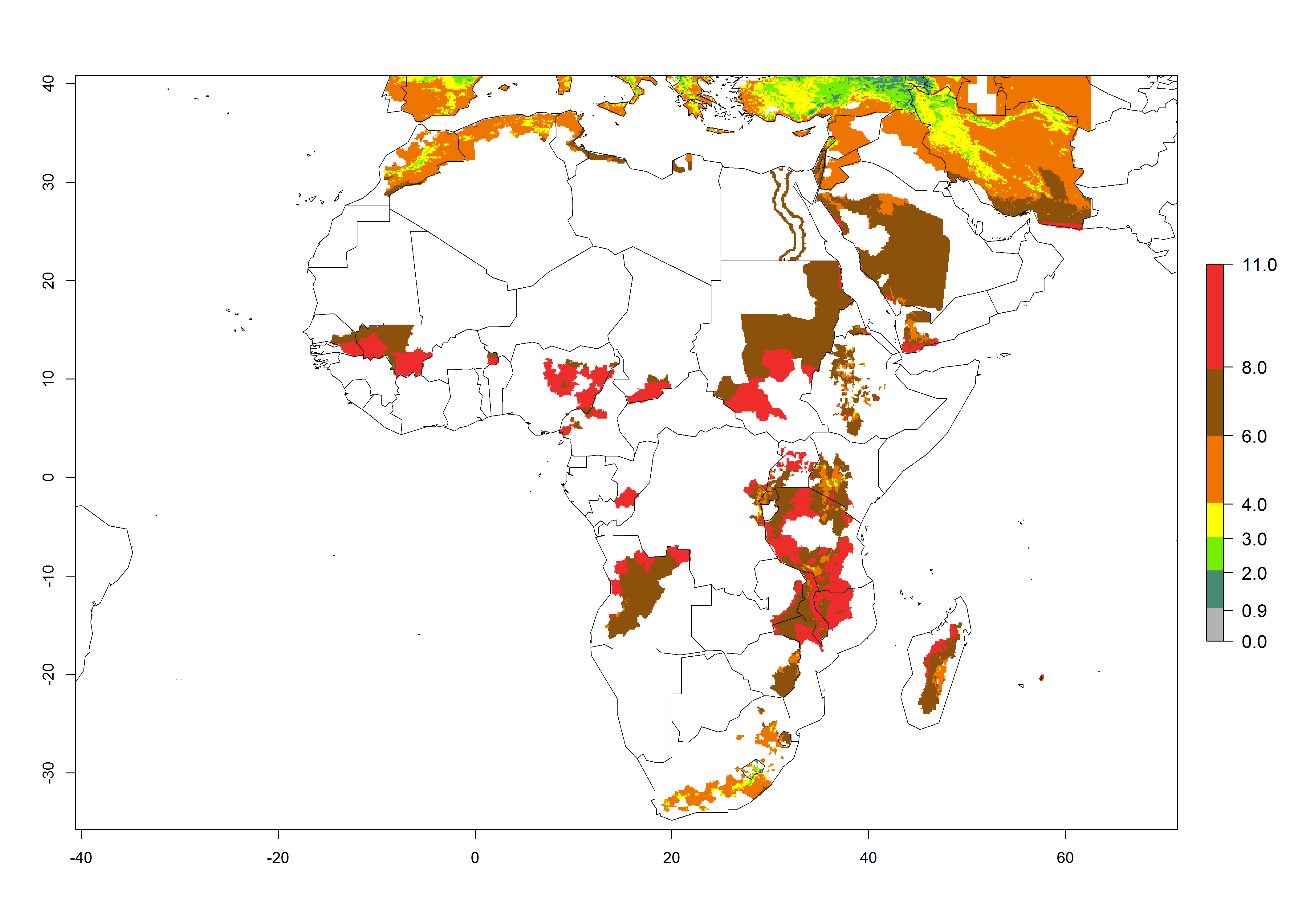 |
 |
| 2050 | 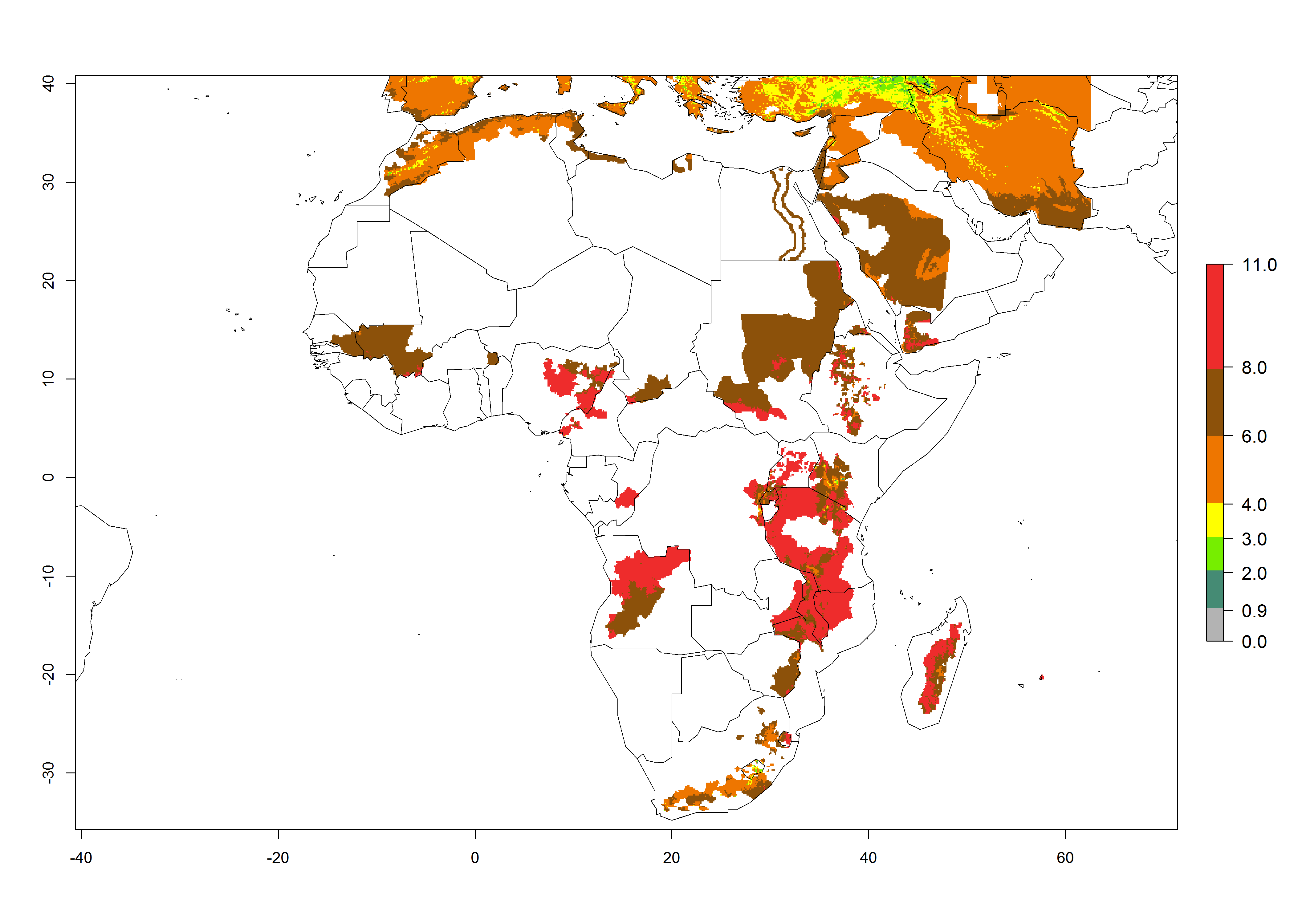 |
 |
| Index change (2000 – 2050) | 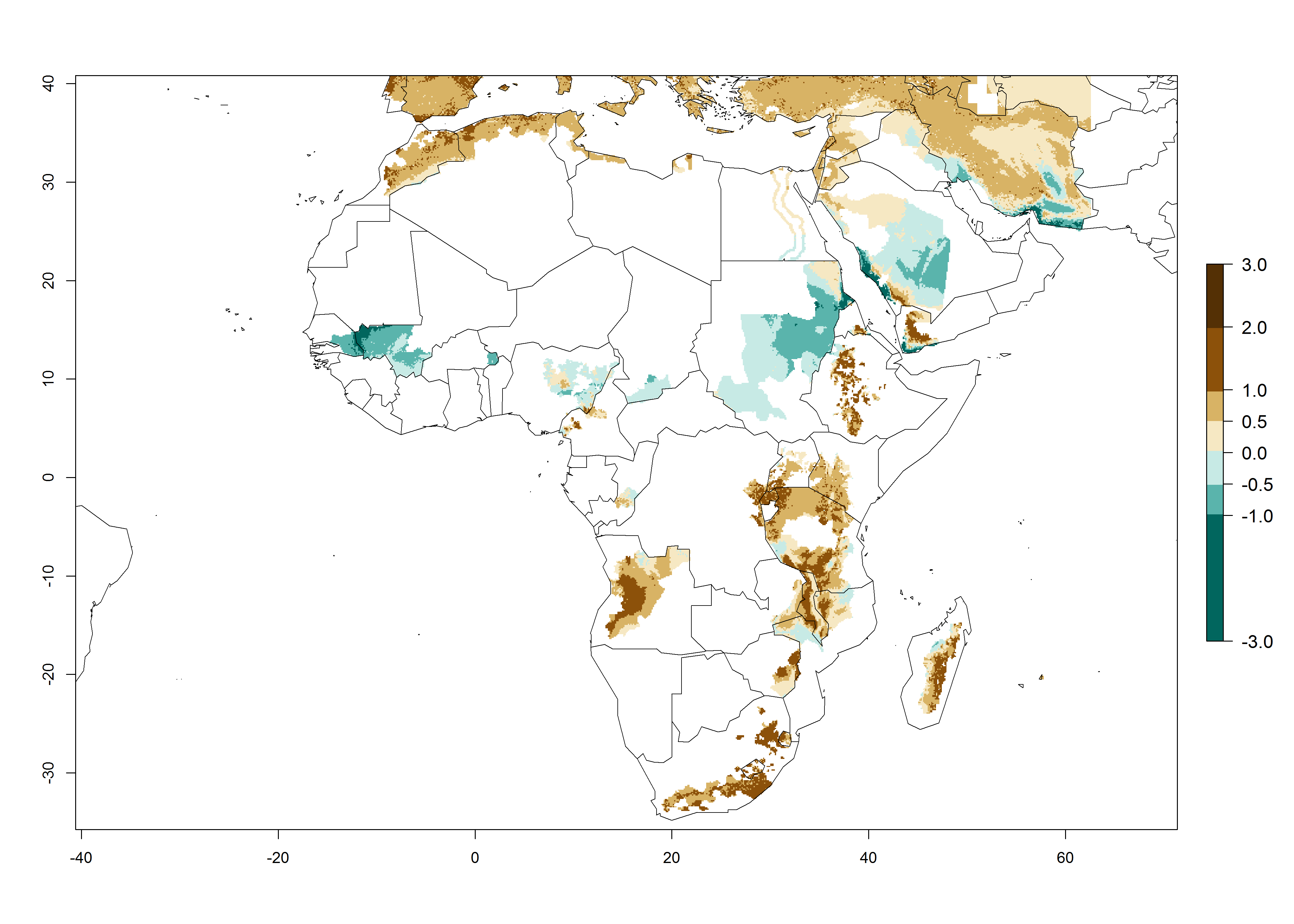 |
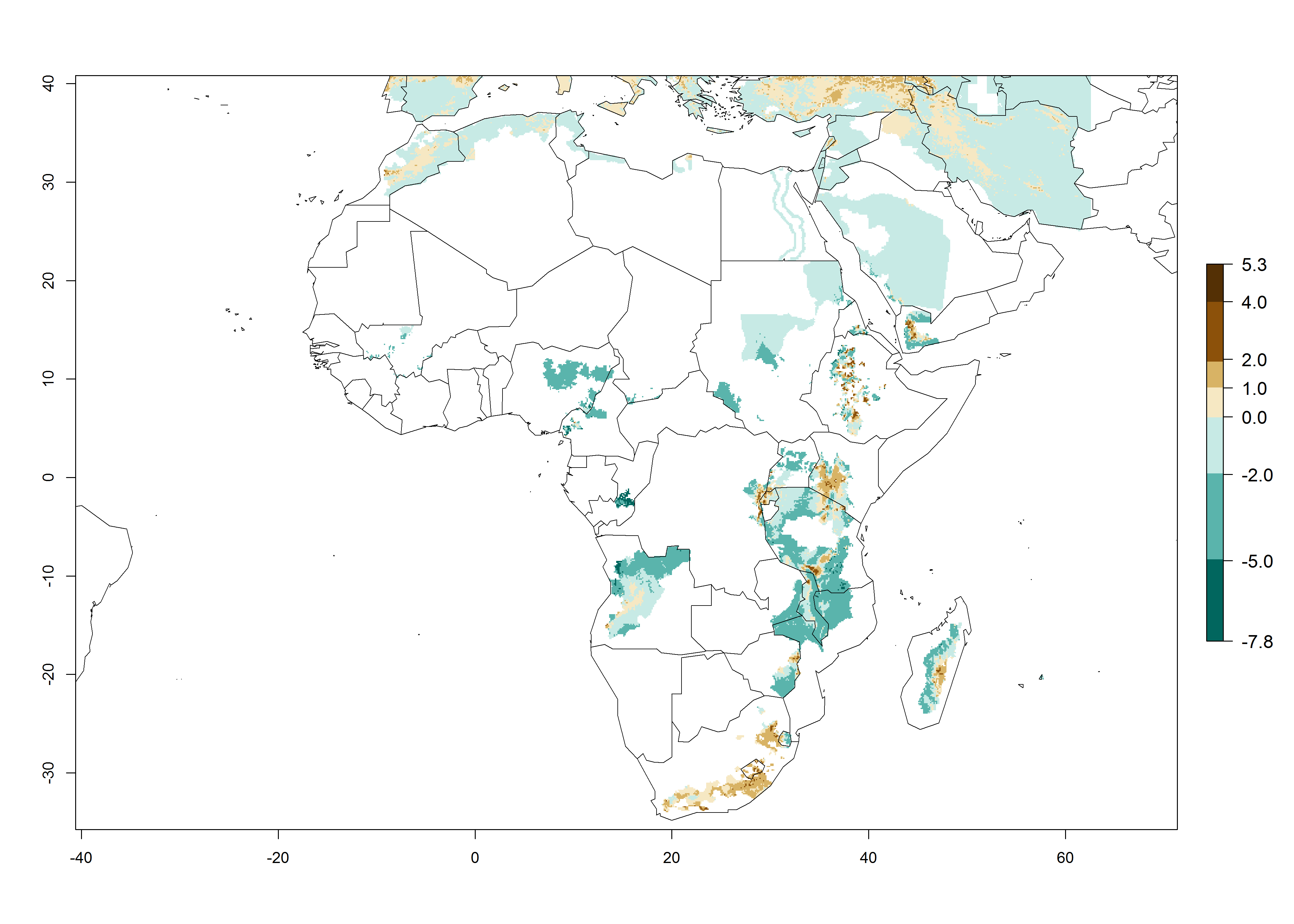 |
Figure 5. Changes in abundance (GI, damage potential) and activity (AI, potential population growth) of the Guatemalan potato tuber moth, Tecia solanivora, in African potato production systems according to model predictions, using the GI (A, D) and the AI (B, E) for the years 2000 and 2050, and the absolute index change (C, F).
Country Risk Maps
Morocco. Potatoes are cultivated from the Atlantic Coast to altitudes of up to 3,000 masl. Establishment of T. solanivora is possible at altitudes up to 1,000 masl. In these regions at risk, 4–6 generations per year might develop and an annual population increase of 104–108 times is estimated (Fig. 6k).
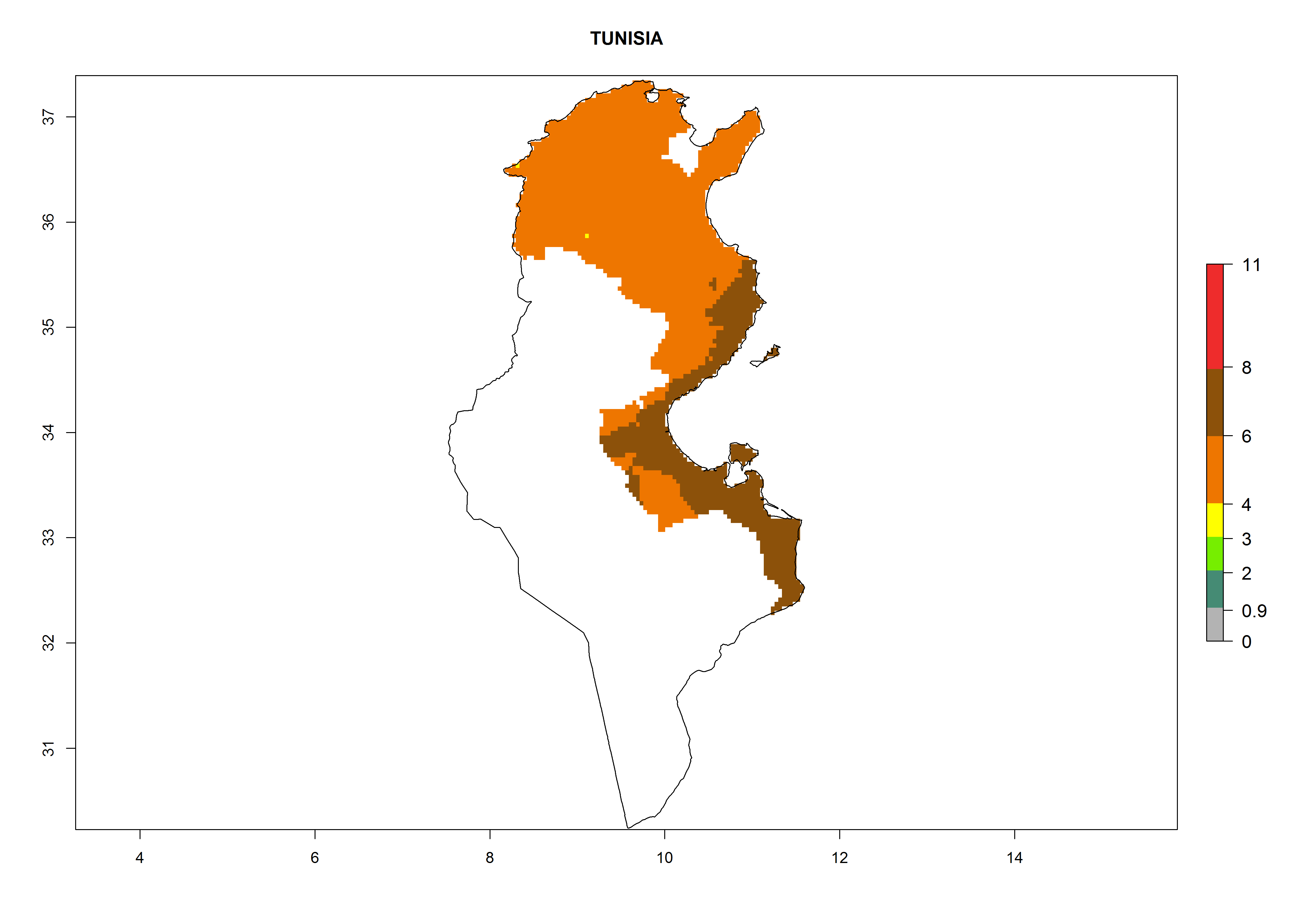
Algeria and Tunisia. Potatoes are cultivated at the Mediterranean Coast from sea level to about 1,200 masl. Potato-growing zones below 600 masl are at risk of T. solanivora establishment. An annual development of 4–6 generations per year with a population increase of 104–108 times is possible (Figs. 6b and 6p).
Egypt. Potatoes are cultivated alongside the Nile. The risk of establishment decreases from the Nile Delta to the frontier with Sudan; establishment is possible in about 40% of the potato-growing zones. Development of 6–8 generations per year and an annual population growth of 104–108 times are estimated (Fig. 6f).
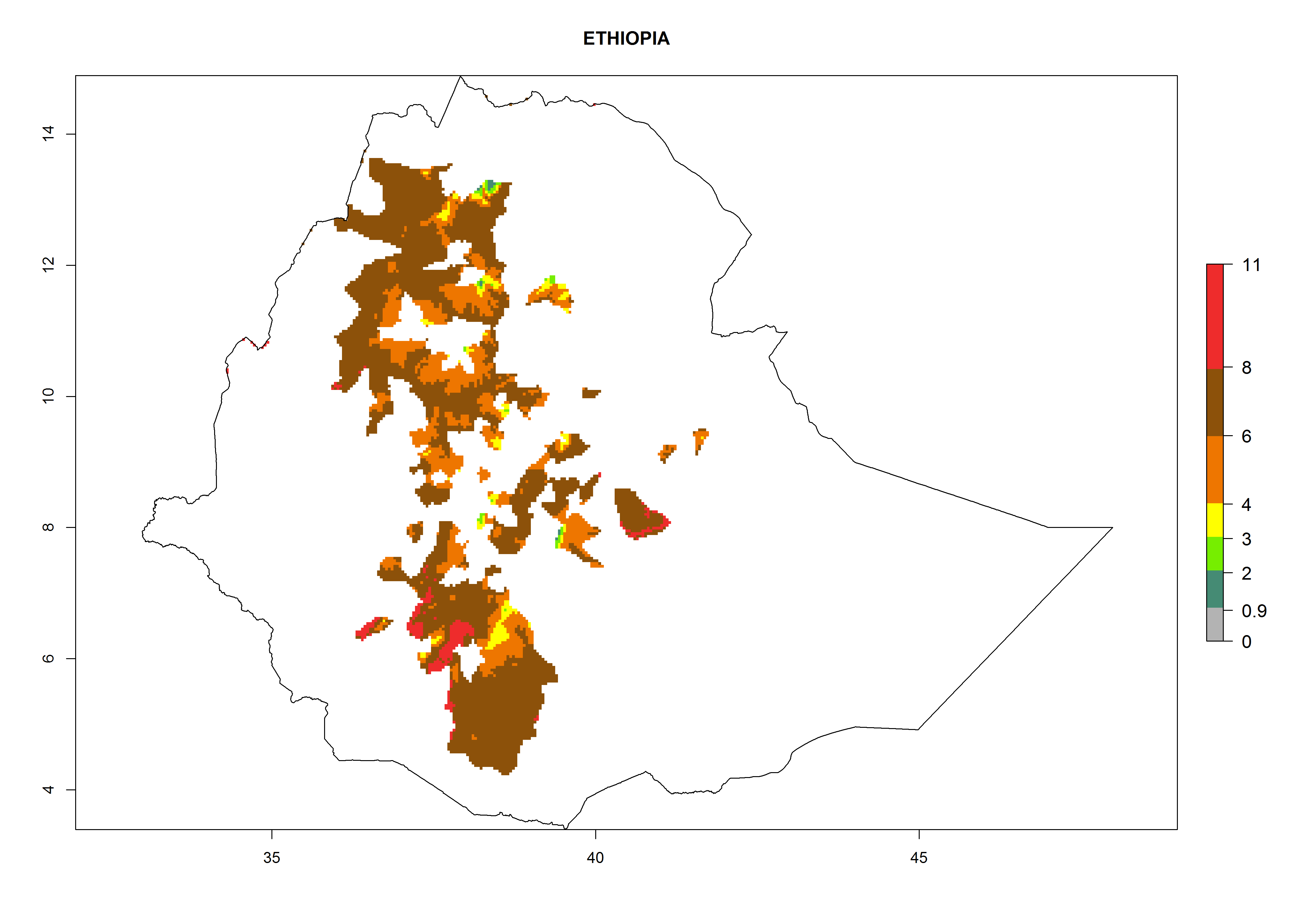
Ethiopia. Potatoes are cultivated in the highlands at 1,500–3,000 masl. T. solanivora establishment is possible in all parts of the potato-cropping area. About 4–8 generations might develop per year and a population increase of 108–1010 per year is estimated (Fig. 6g).
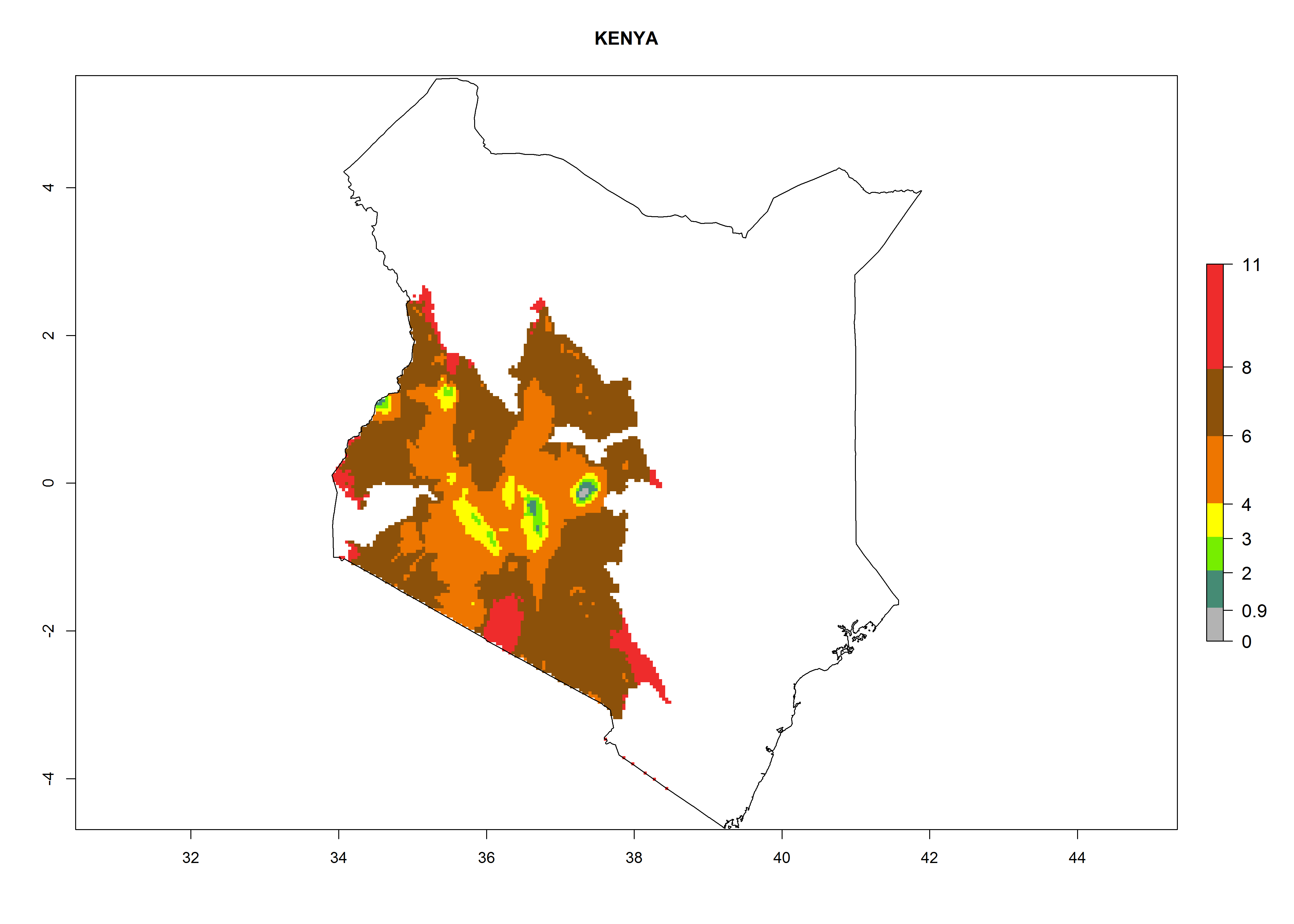
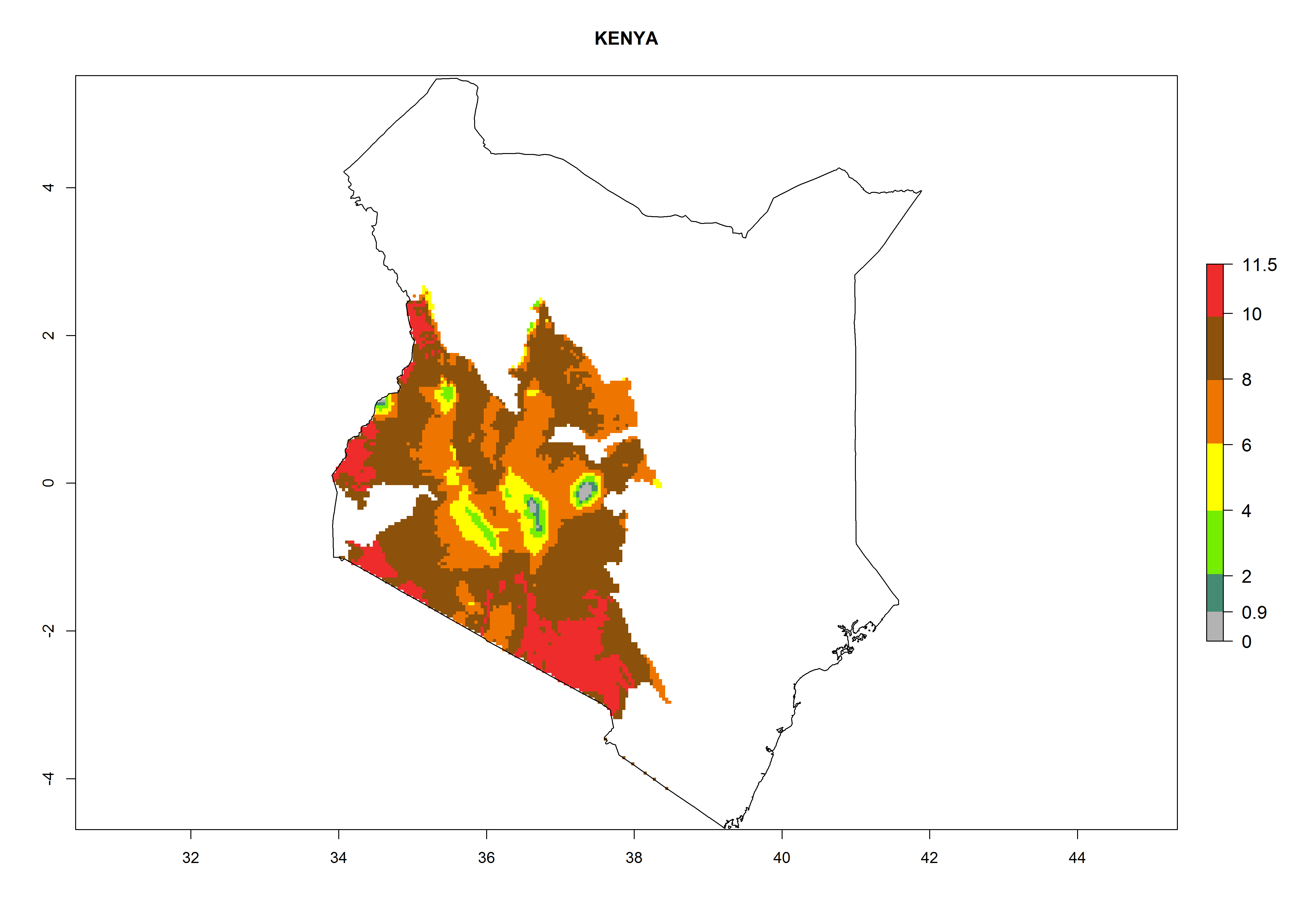
Kenya. Potatoes are cultivated in the southwest mountain area at 1,000–3,000 masl. Establishment is possible for these zones and 6–10 generations per year might develop. An annual population increase of 108–1011.5 is estimated (Fig. 6h).
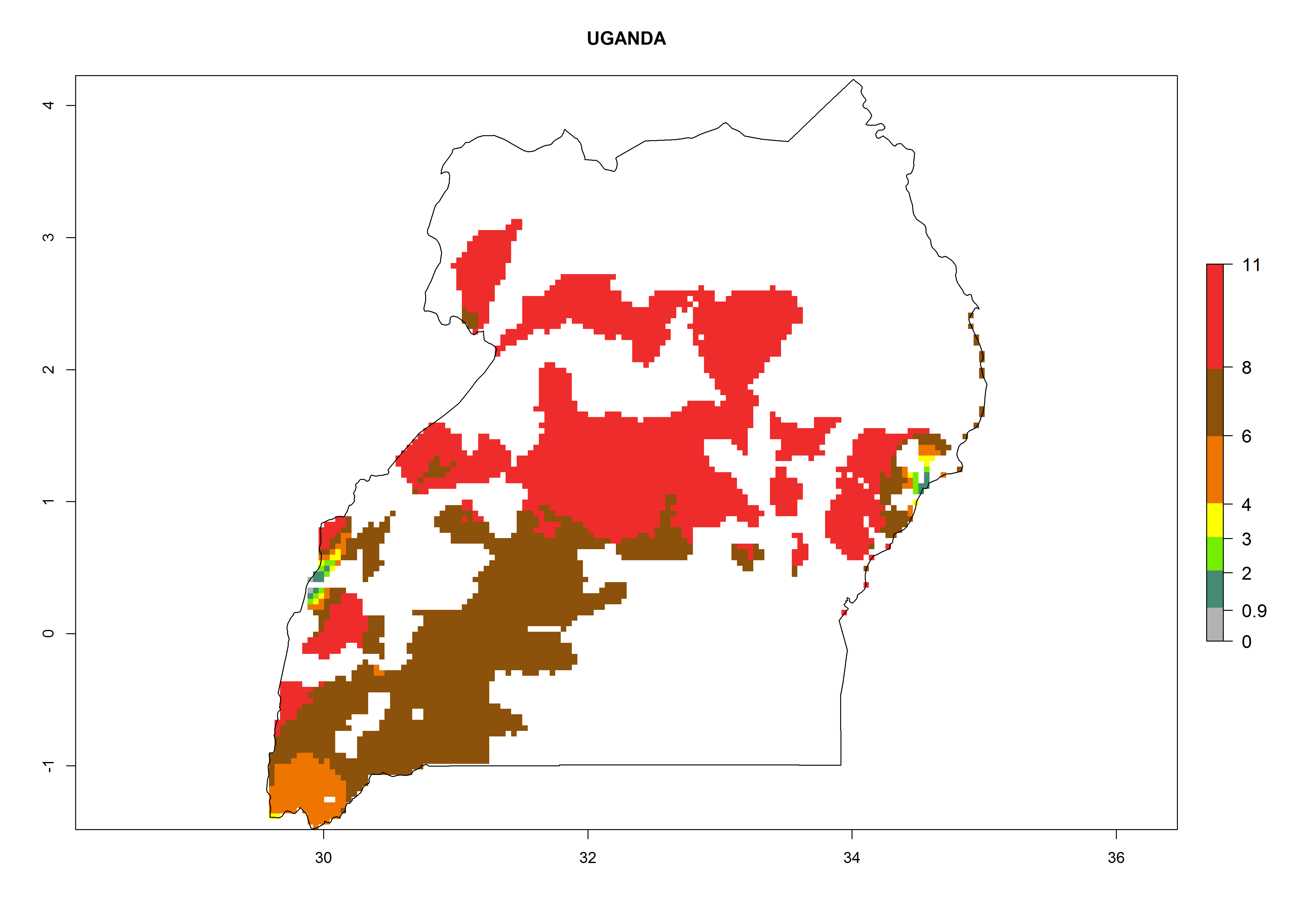
Uganda. Potatoes are cultivated from 1,000 masl in the north to 2,200 masl in the southwest. An ERI>0.95 is indicated for the complete potato cultivation area. With increasing altitude, the number of generations is reduced from 10.5 to 6, but at the same time the population growth potential increases from 108 to 1011.5 per year (Fig. 6q).
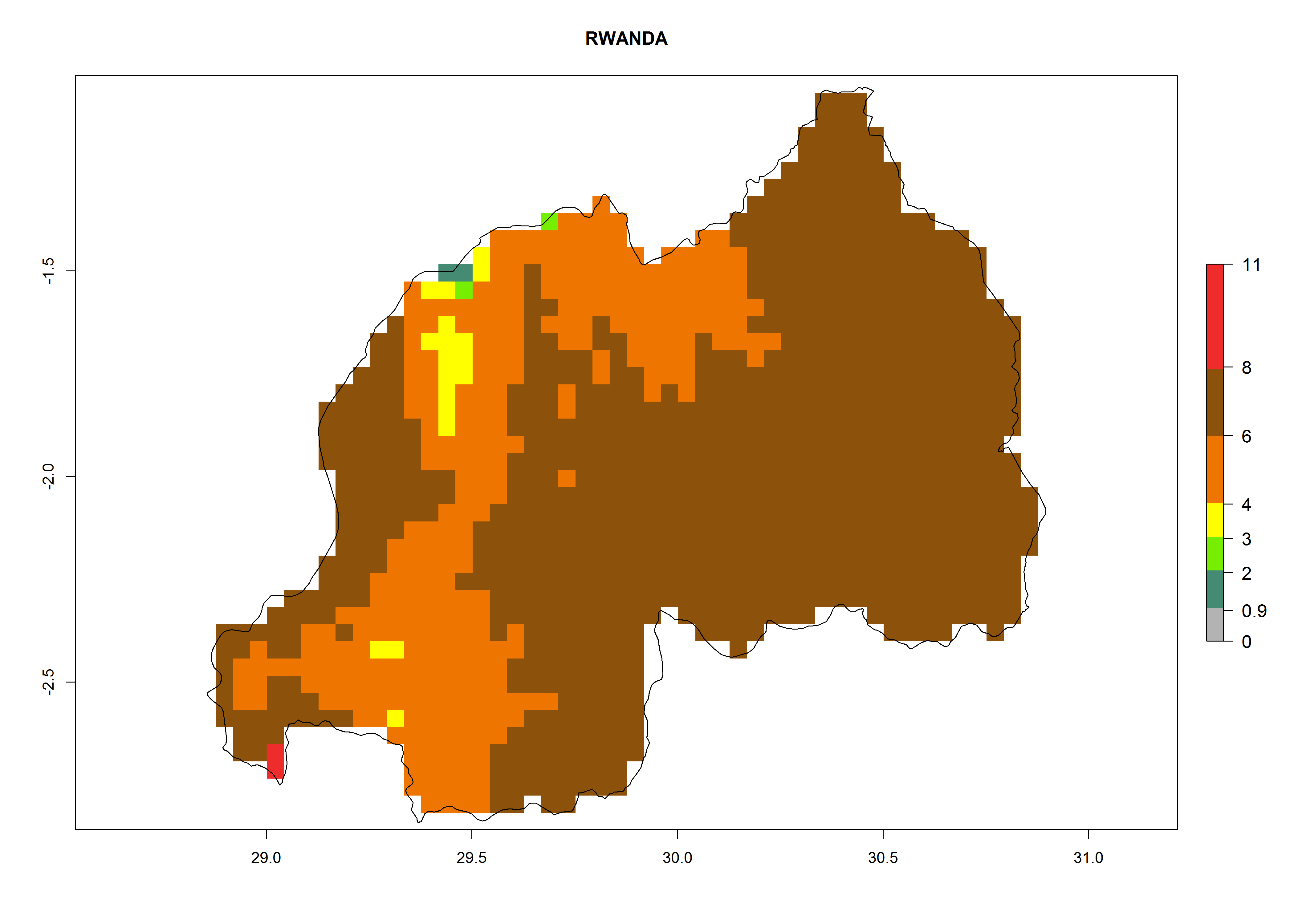
Rwanda. Potatoes are cultivated in all parts of the country and temperature conditions are favorable for T. solanivora establishment in all regions. For the mountain region in the west, with elevations up to 3,000 masl, 4–6 generations per year are predicted. For the eastern plateau, at about 1,500 masl, 6–8 generations might develop. The potential population increase is also higher for the eastern part of the country (Fig. 6m).
DR Congo. Potatoes are only cultivated in small parts of the eastern mountain region close to the frontiers with Uganda and Rwanda at an altitude of 1,000–3,000 masl. Establishment in this region is possible, and the potential population growth is high: with an AI>10 for most of the area and 4–11 generations per year (Fig. 6e).
Congo. Potatoes are cultivated on a plateau at about 500–1,000 masl in the middle of the country. Establishment is possible in all parts the potato-growing zone, and an annual population increase of 106–1010 is estimated. About 8–10 generations per year might develop (Fig. 6d).
Cameroon. Potatoes are cultivated mainly in the western highlands at up to 1,500 masl; establishment is possible in all potato-growing zones. Per year, 6–11 generations might develop with an estimated population increase of 108–1011.5 for most parts (Fig. 6c).
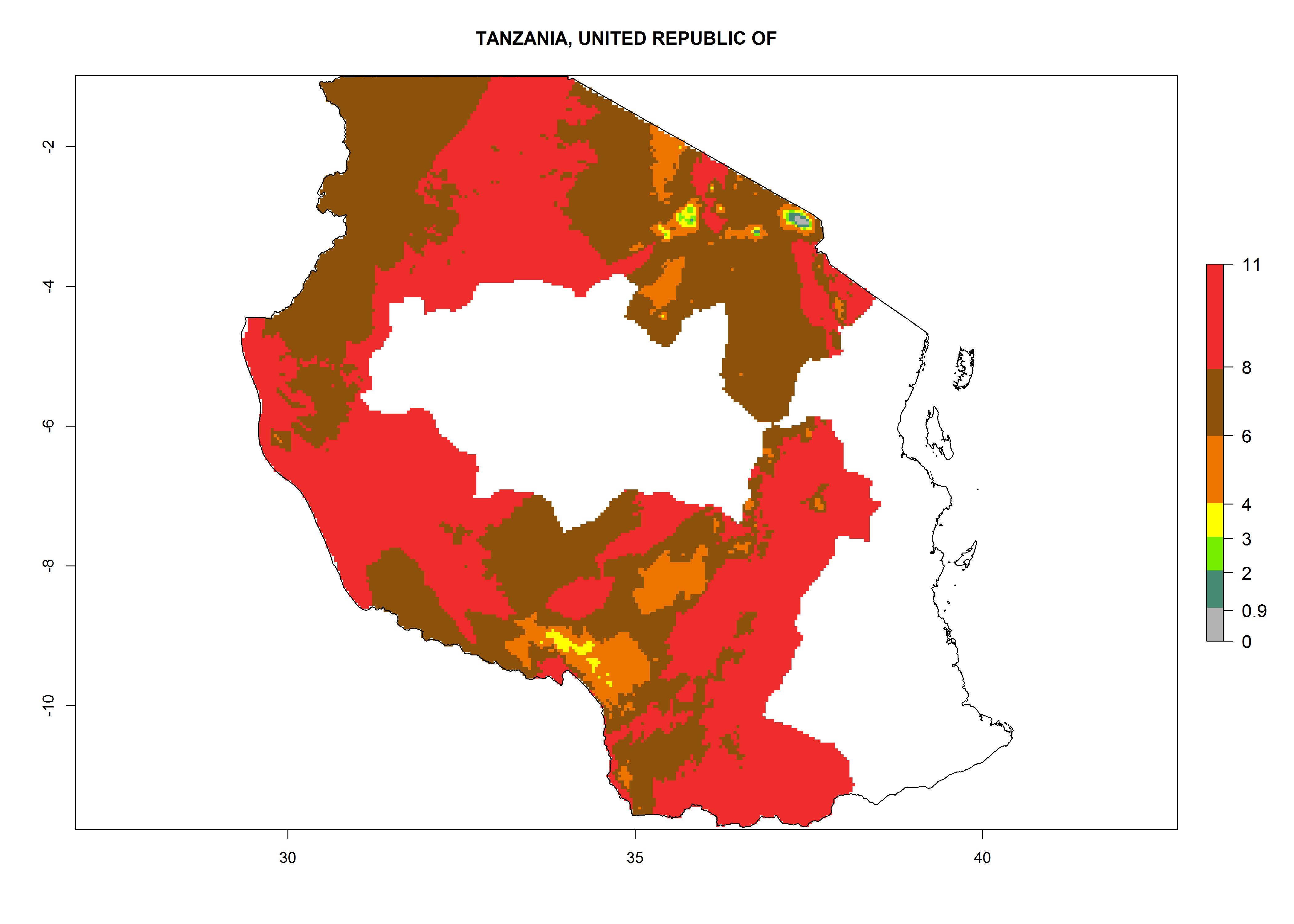
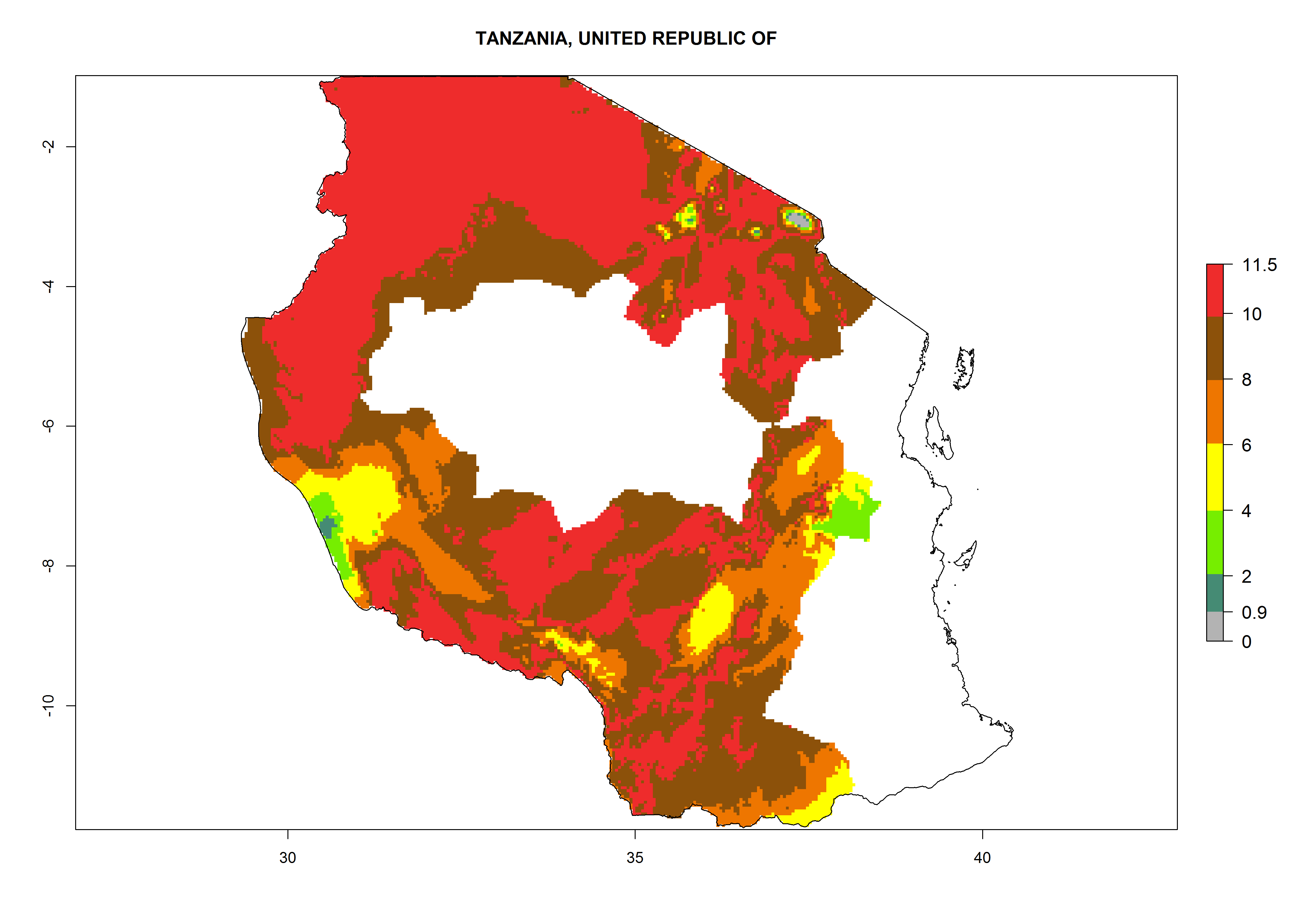
Tanzania. Establishment is possible in most parts of the potato-growing zones. Six or more generations per year might develop in all potato-growing zones except in areas above 2,000 masl, where 4–6 generations are expected. A potential annual population increase of 108–1011.5 is estimated for most potato-growing zones (Fig. 6p).
Malawi. Establishment is possible in the northern and central areas; 6–8 generations per year might develop (Lake Malawi is in the eastern part of the country, where a higher number of generations is indicated on the map, Fig. 6j) and an annual population increase of 108–1011.5 is estimated.
Angola. Potatoes are cultivated in the highlands at 1,000–2000 masl. Establishment is possible in all potato-growing areas. Per year 6–11 generations might develop and an annual population increase of 108–1011.5 times is estimated (Fig. 6a).
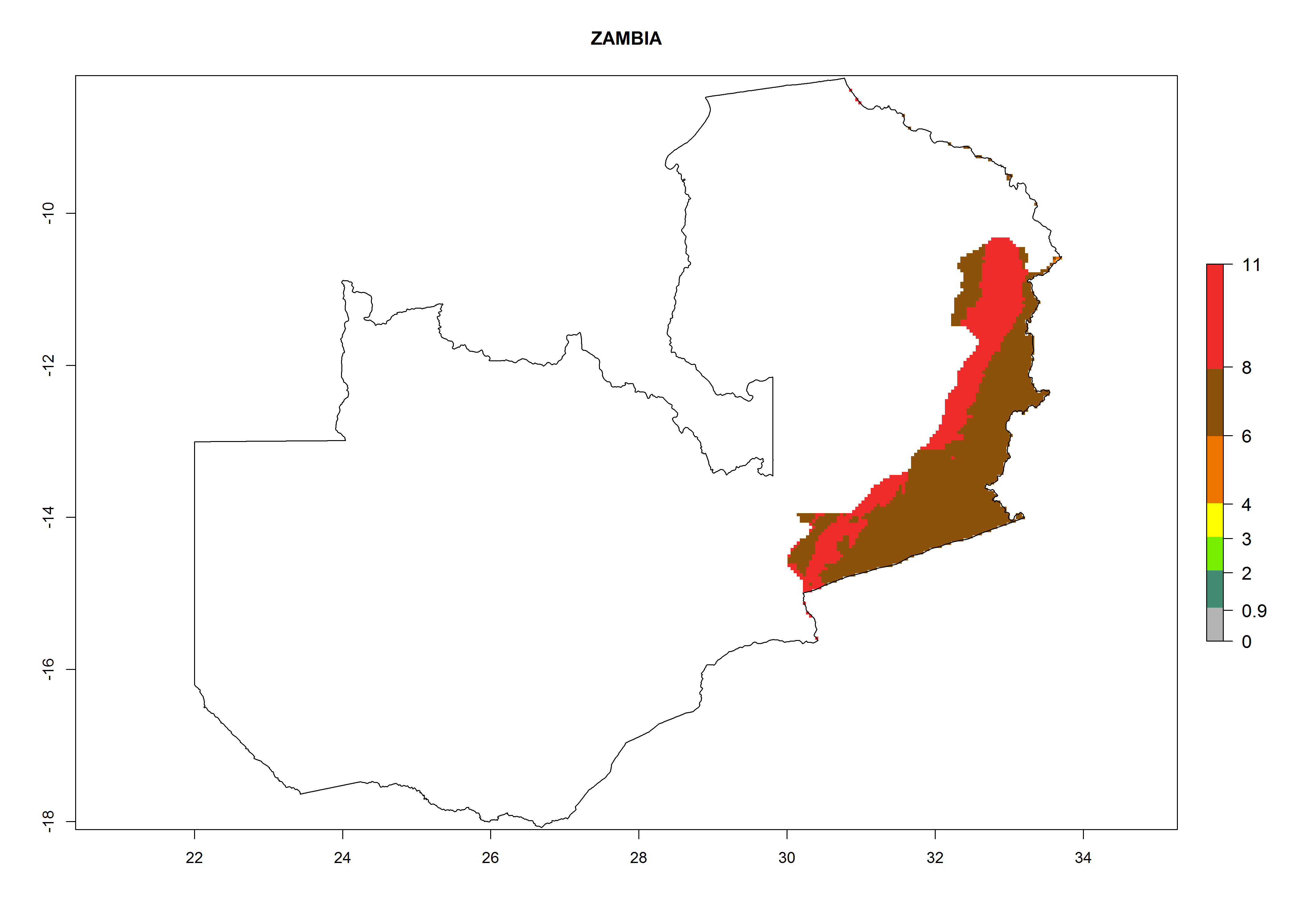
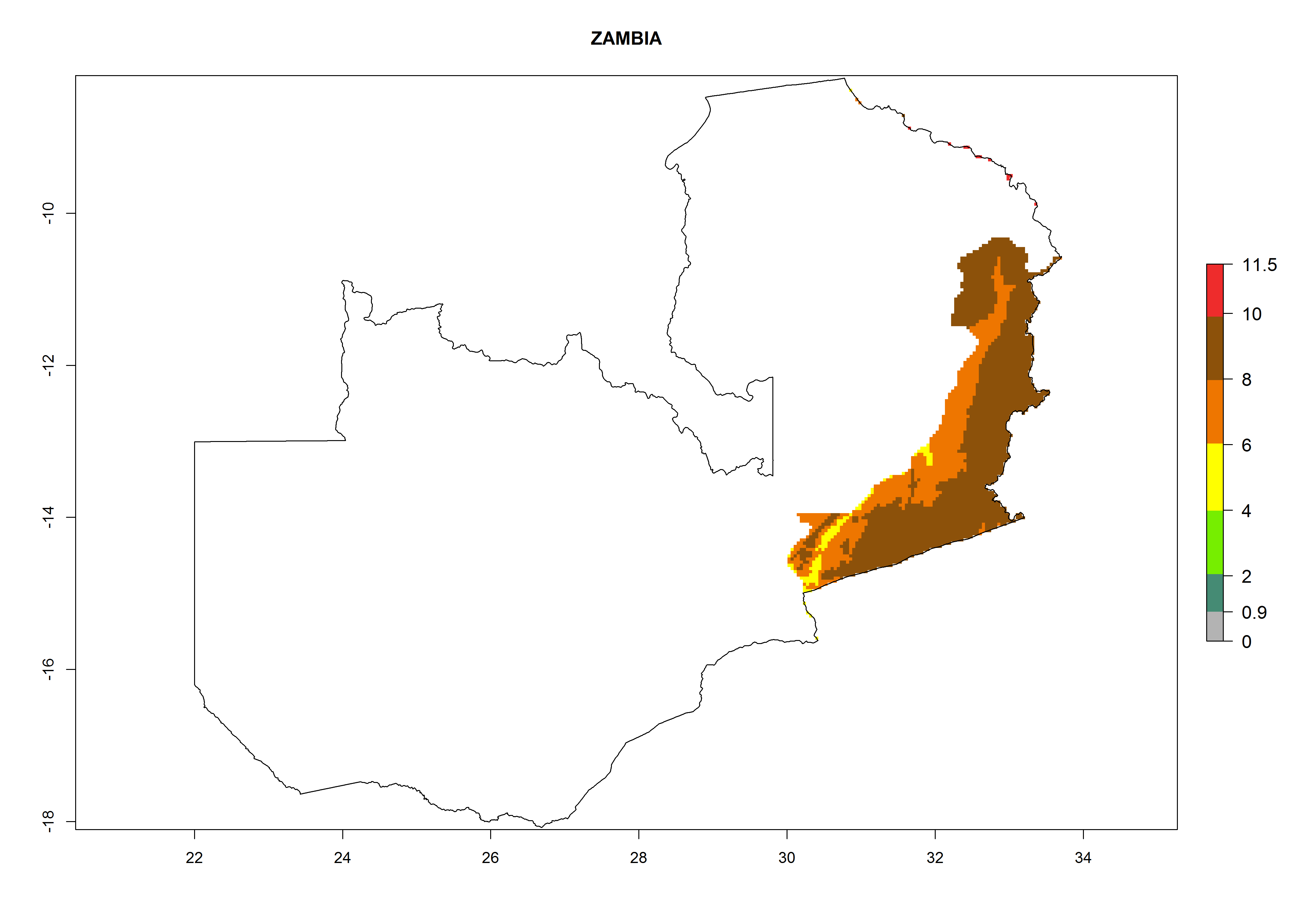
Zambia. Potatoes are cultivated in the eastern part of the country alongside the frontier with Malawi at 500–1,000 masl. Development of 6–11 generations per year and an annual population growth of 106–1010 times is estimated (Fig. 6r).
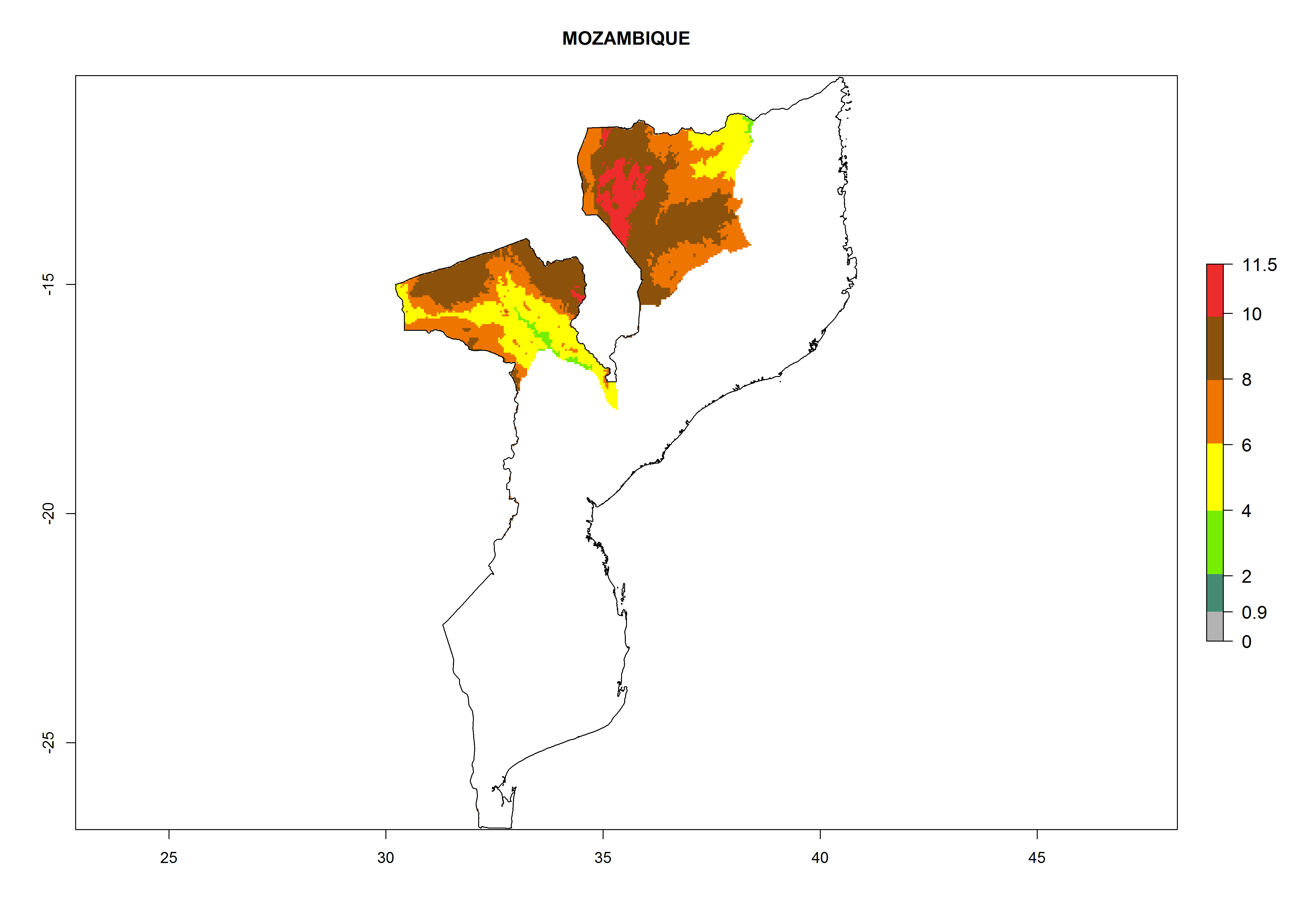
Mozambique. Potatoes are cultivated in the highlands in the northwest. T. solanivora establishment is possible above 600 masl. Within one year 6–8 generations might develop and the population might increase 108–1011.5 times (Fig. 6l).
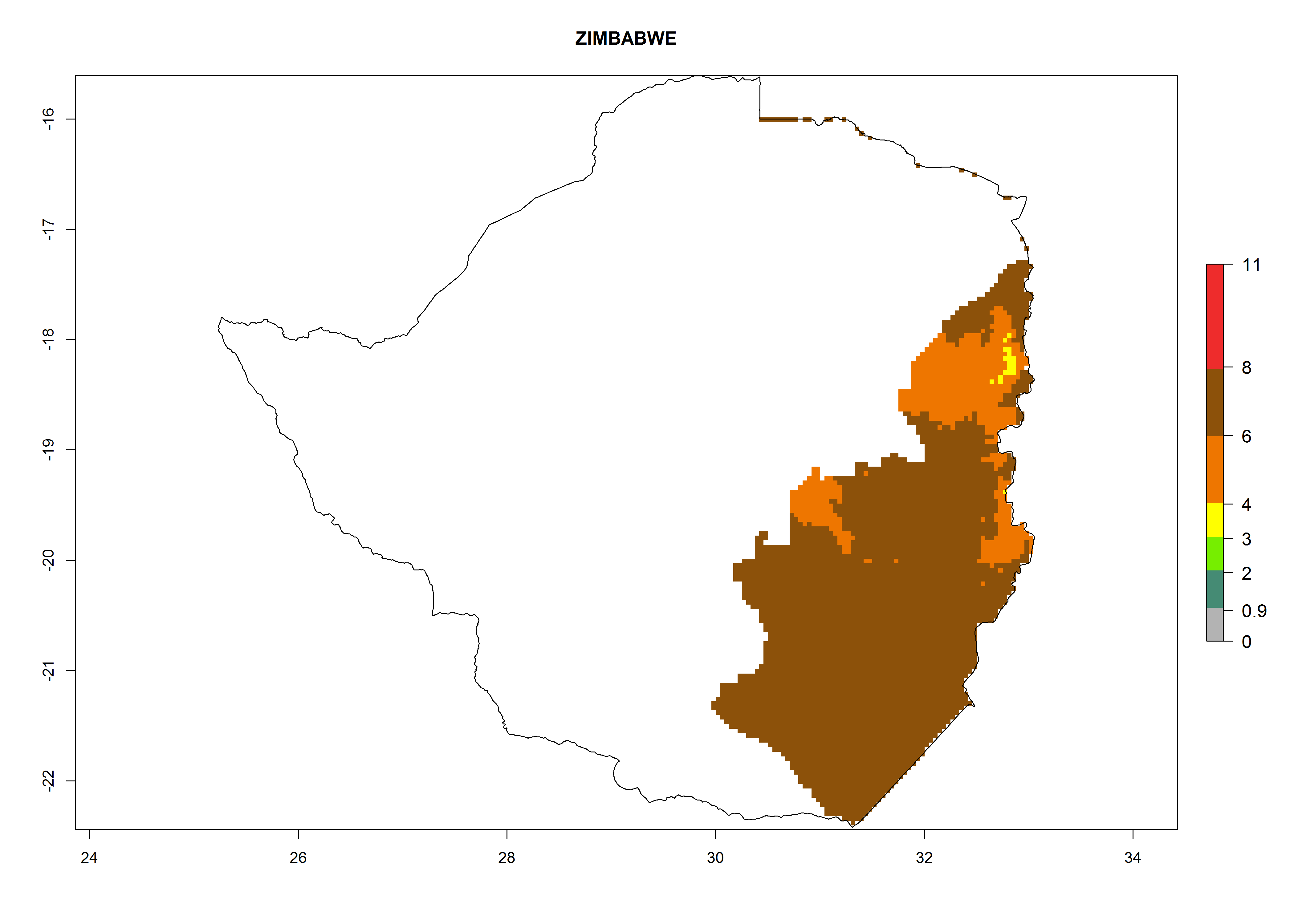
Zimbabwe. Potatoes are cultivated in the east of the country close to the border of Mozambique, where T. solanivora establishment is possible. The northern part of the potato-growing zone is located at 1,000–2,000 masl. There, 4–8 generations might develop per year. The potential annual population increase is 108–1010 times. The southern part of the potato-growing zone is located at below 1,000 masl, where 6–8 generations might develop per year with an annual population increase rate of 106–108 (Fig. 6s).
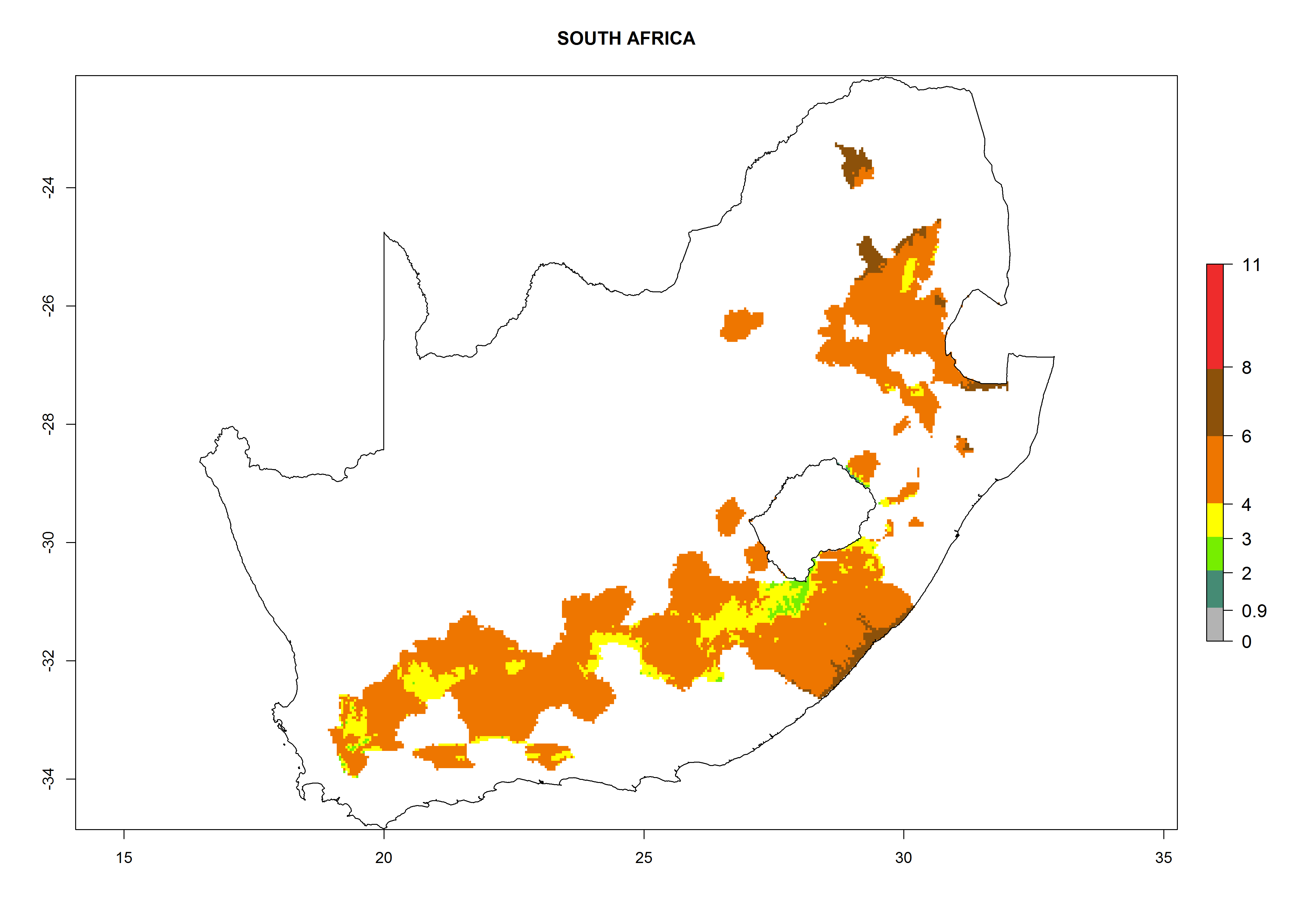
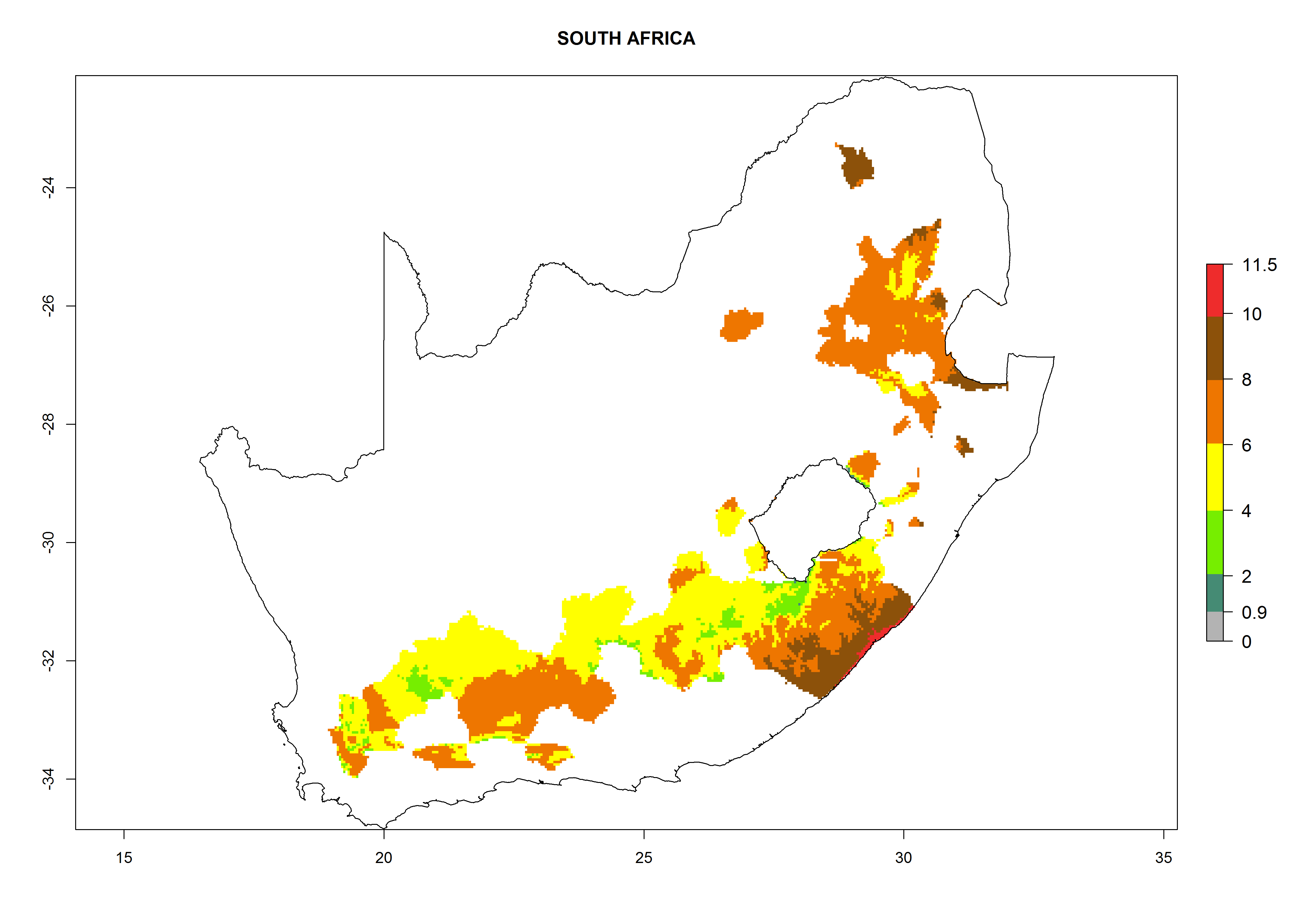
South Africa. Establishment is possible in most parts of the potato-growing zones. About 4–6 generations per year might develop and an annual population increase of 104–108 is possible. In the west of the country, where the climate is milder during winter, this potential for population growth could be as high as 1010 (Fig. 6n).
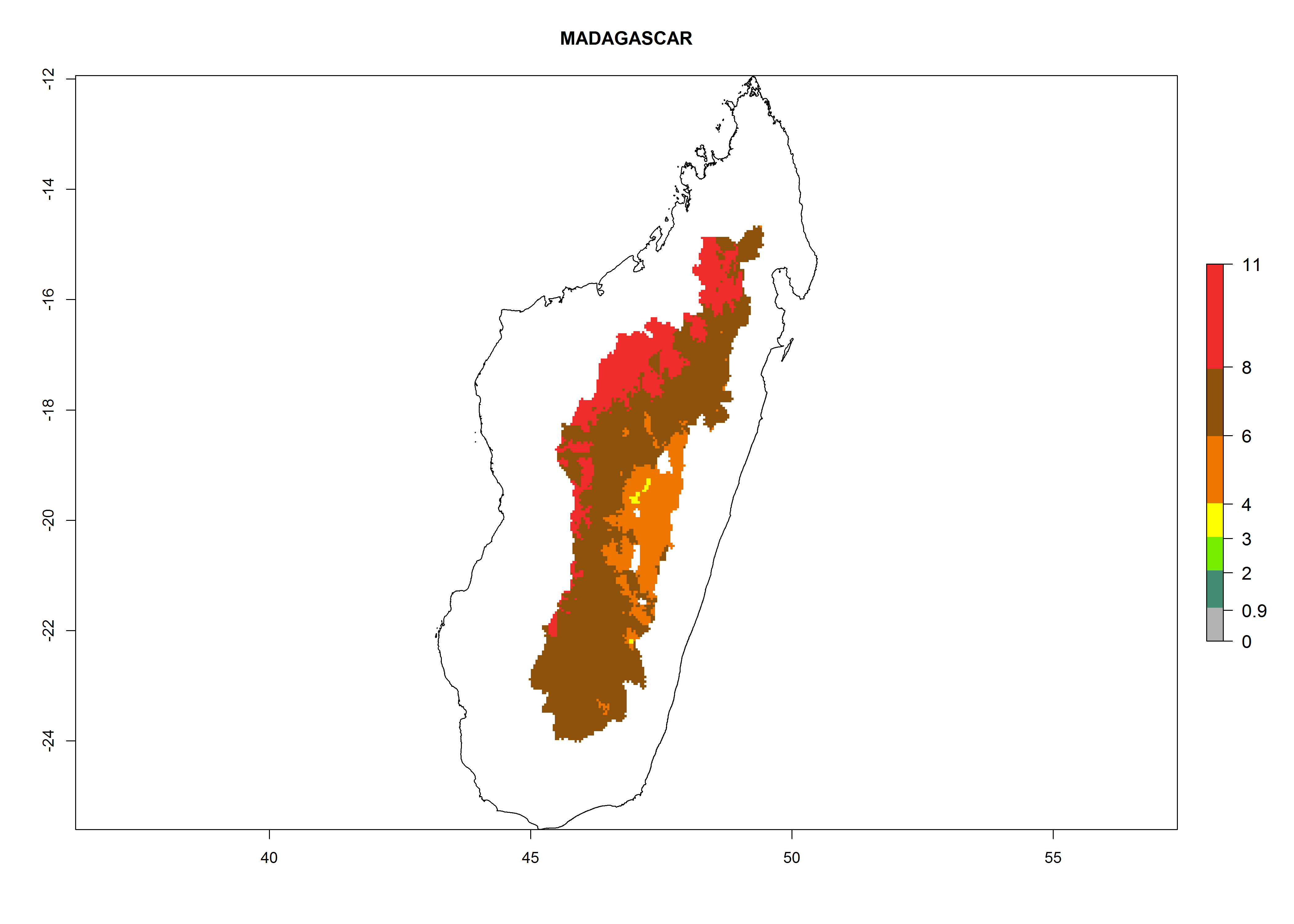
Madagascar. Potatoes are cultivated in the mountains at altitudes of 1,000–2,000 masl. Establishment is possible in most parts of the potato-growing zone; 4–8 generations per year might develop. An annual population growth of 106–1010 is possible (Fig. 6i).
| ERI | GI | AI |
| a) Angola
|
 |
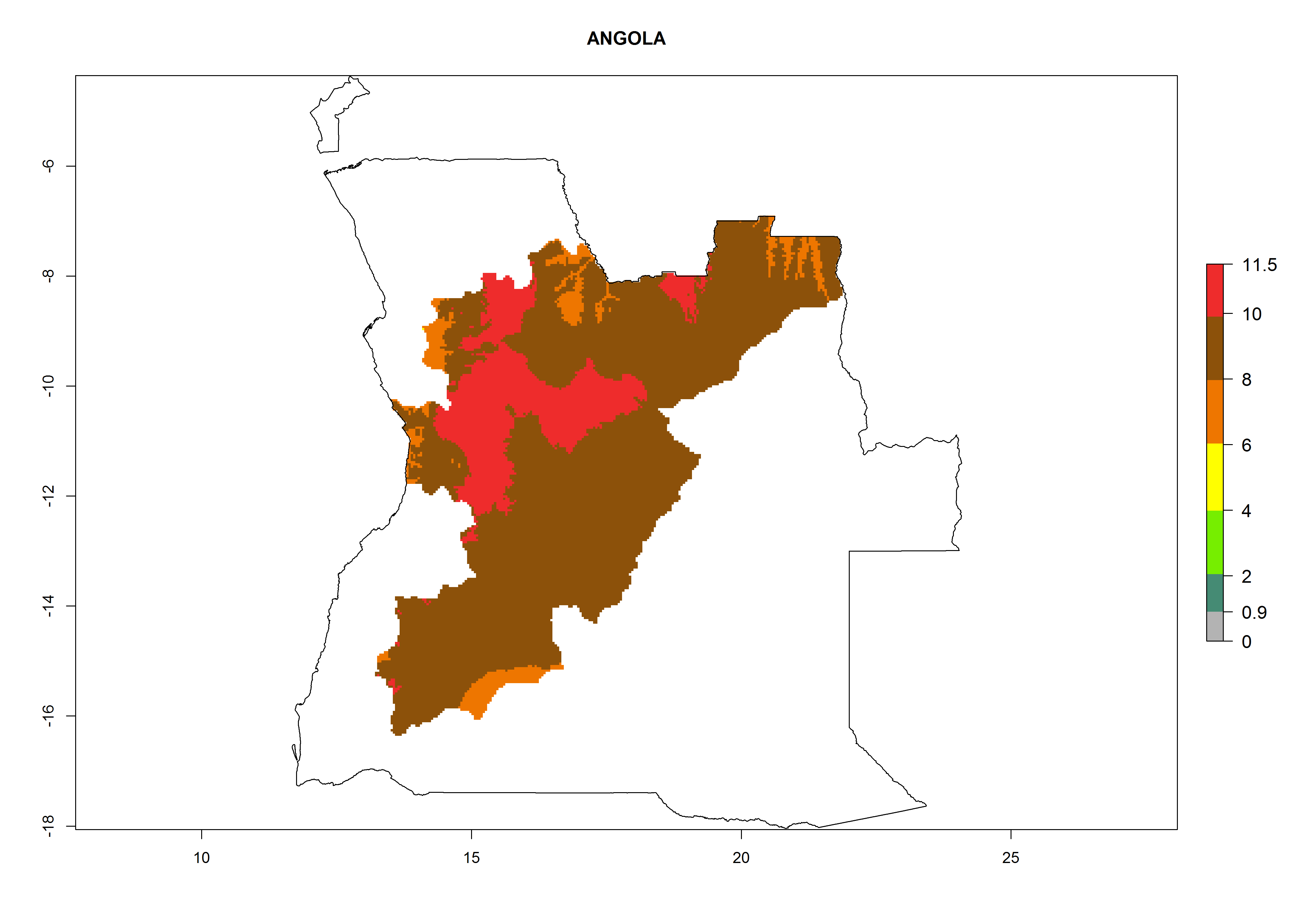 |
| b) Algeria
|
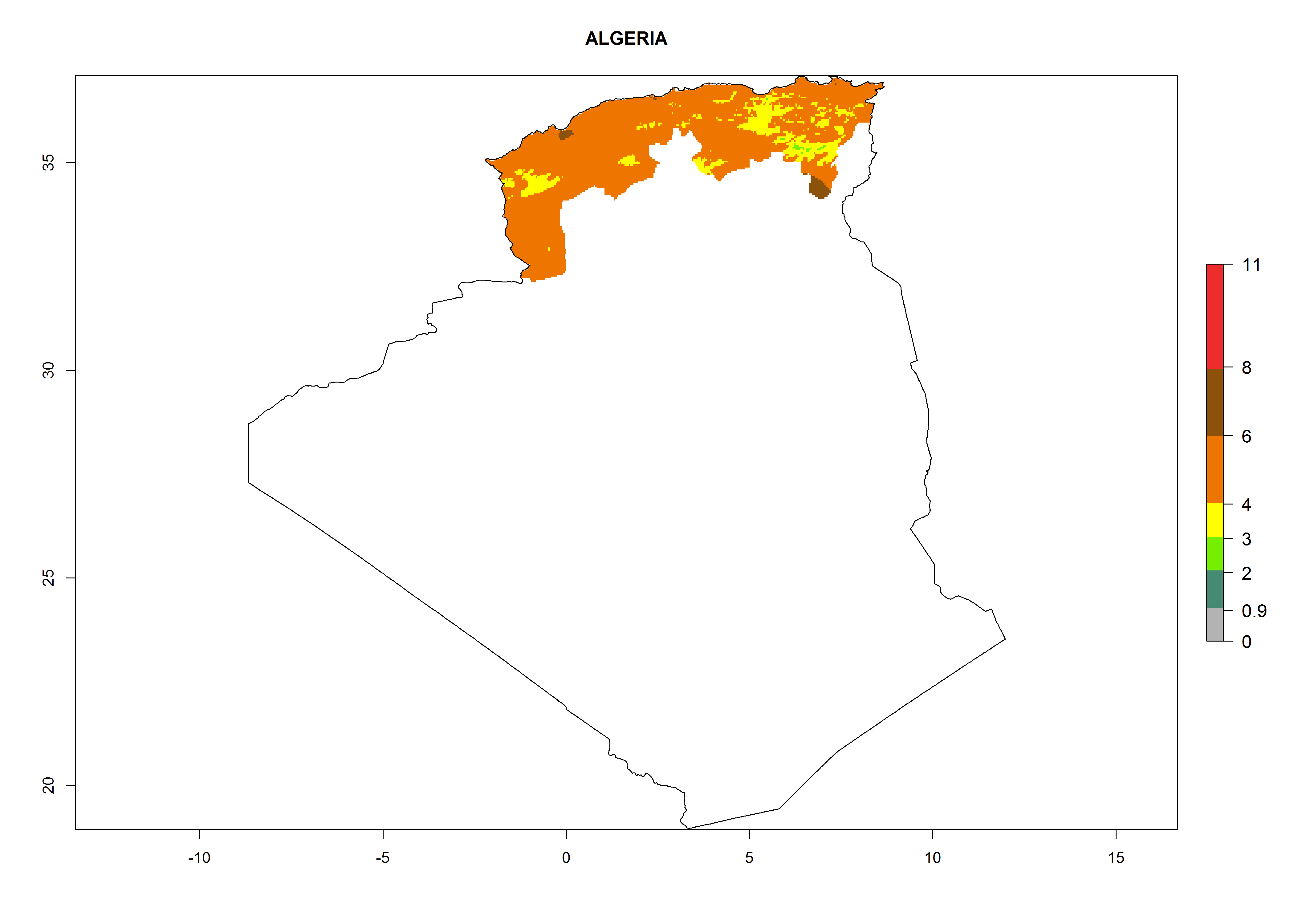 |
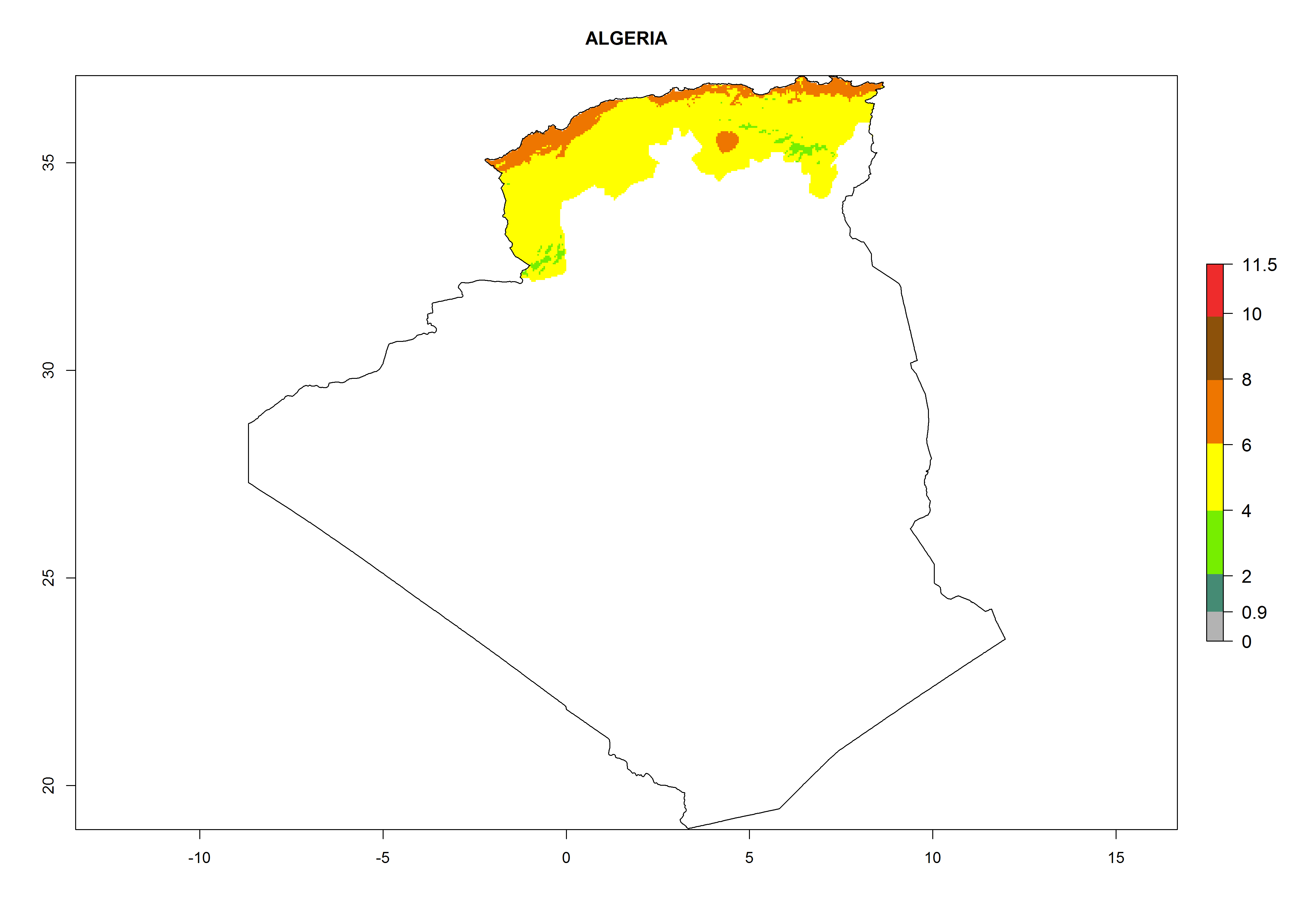 |
| c) Cameroon
|
 |
 |
| d) Congo
|
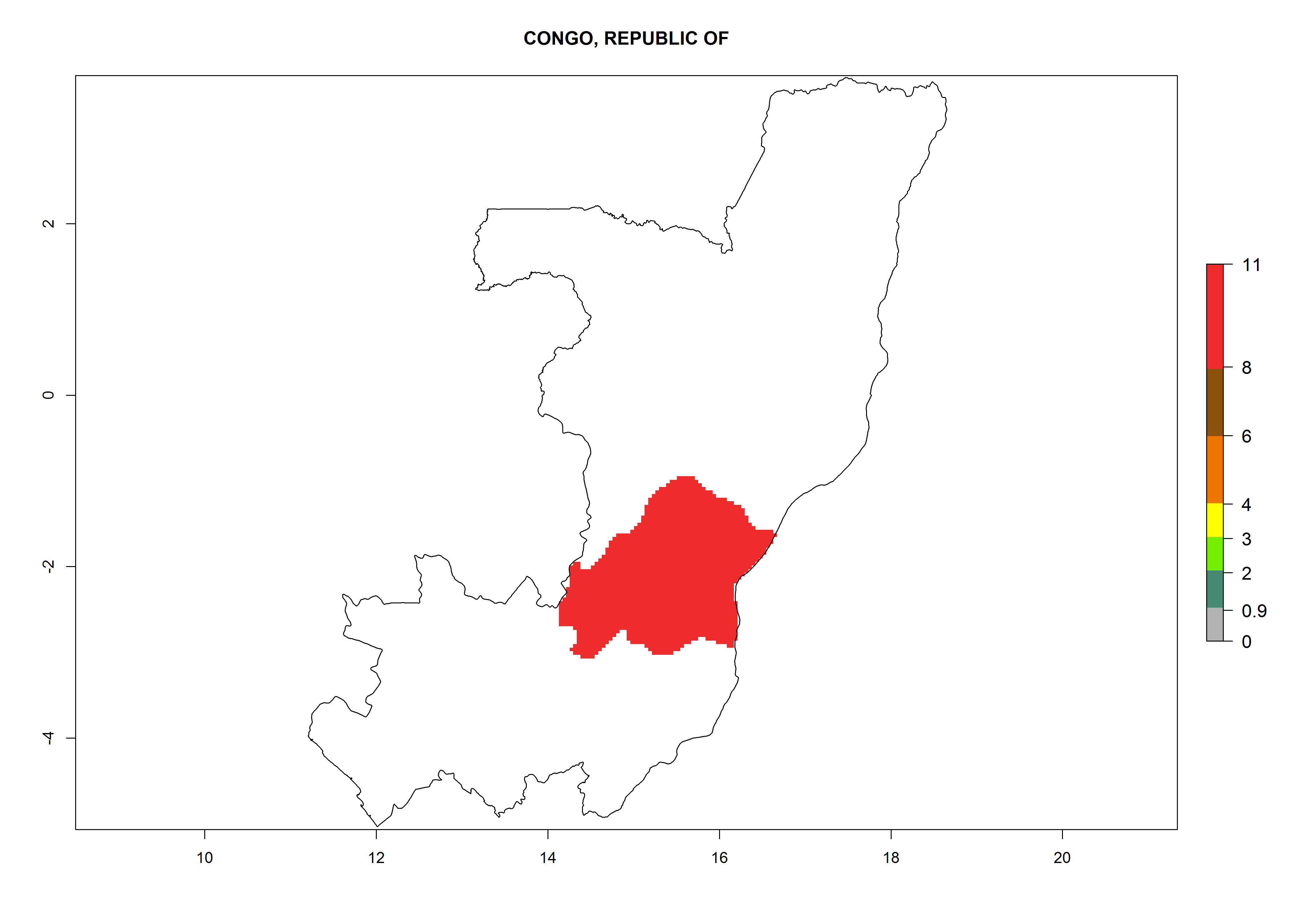 |
 |
| e) DRC
|
 |
 |
| f) Egypt
|
 |
 |
| g) Ethiopia
|
 |
 |
| h) Kenya
|
 |
 |
| i) Madagascar
|
 |
 |
| j) Malawi
|
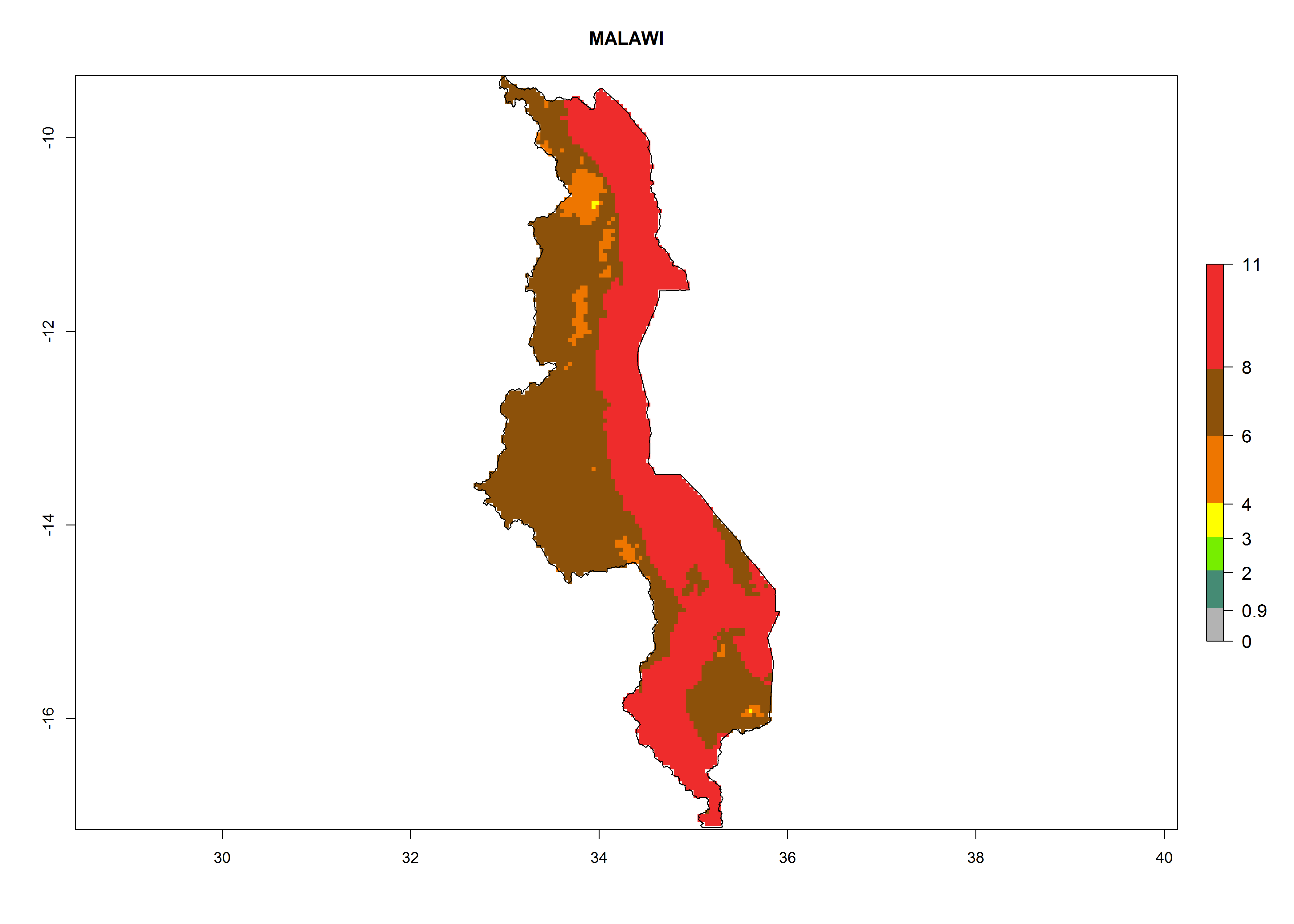 |
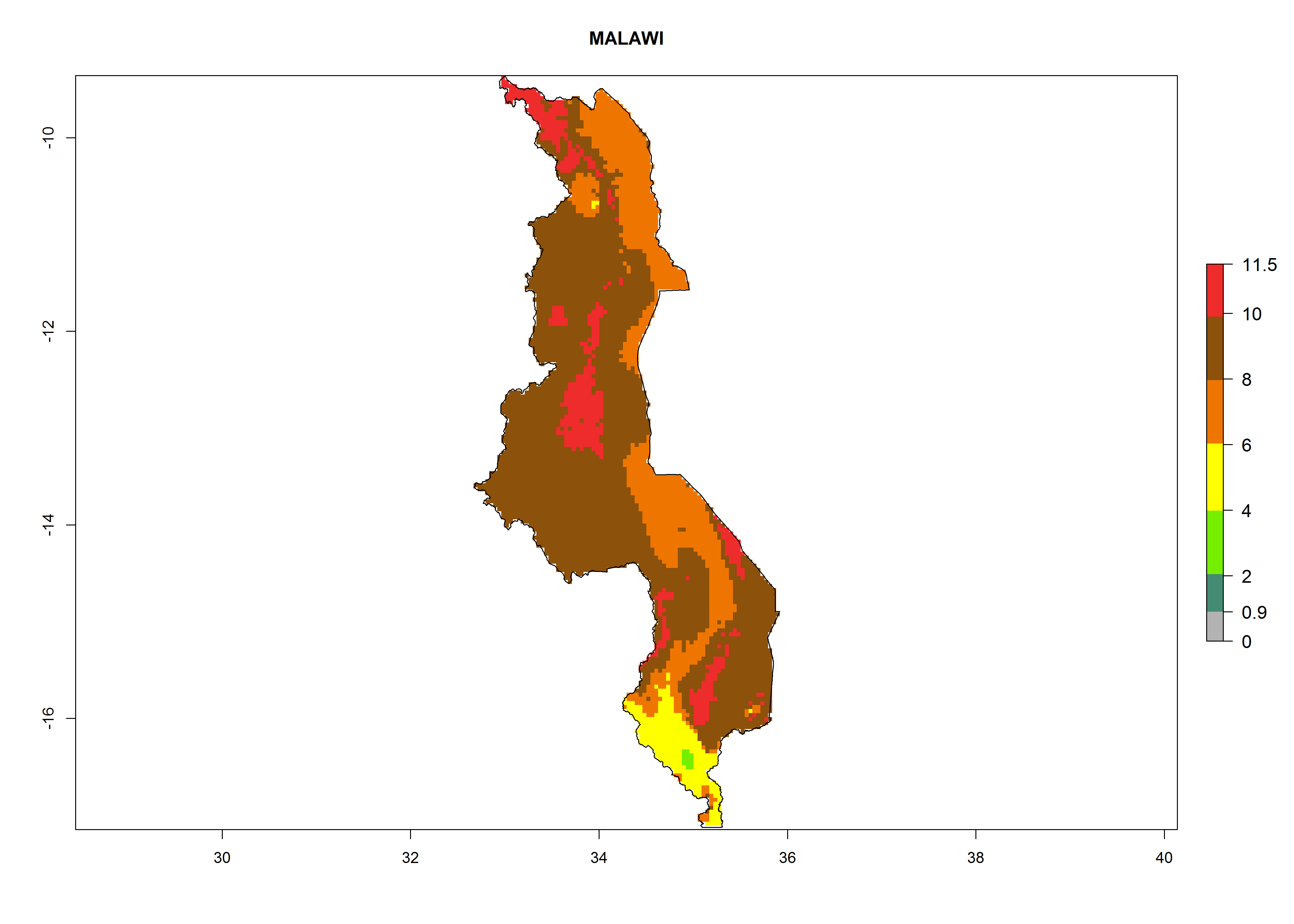 |
| k) Morocco
|
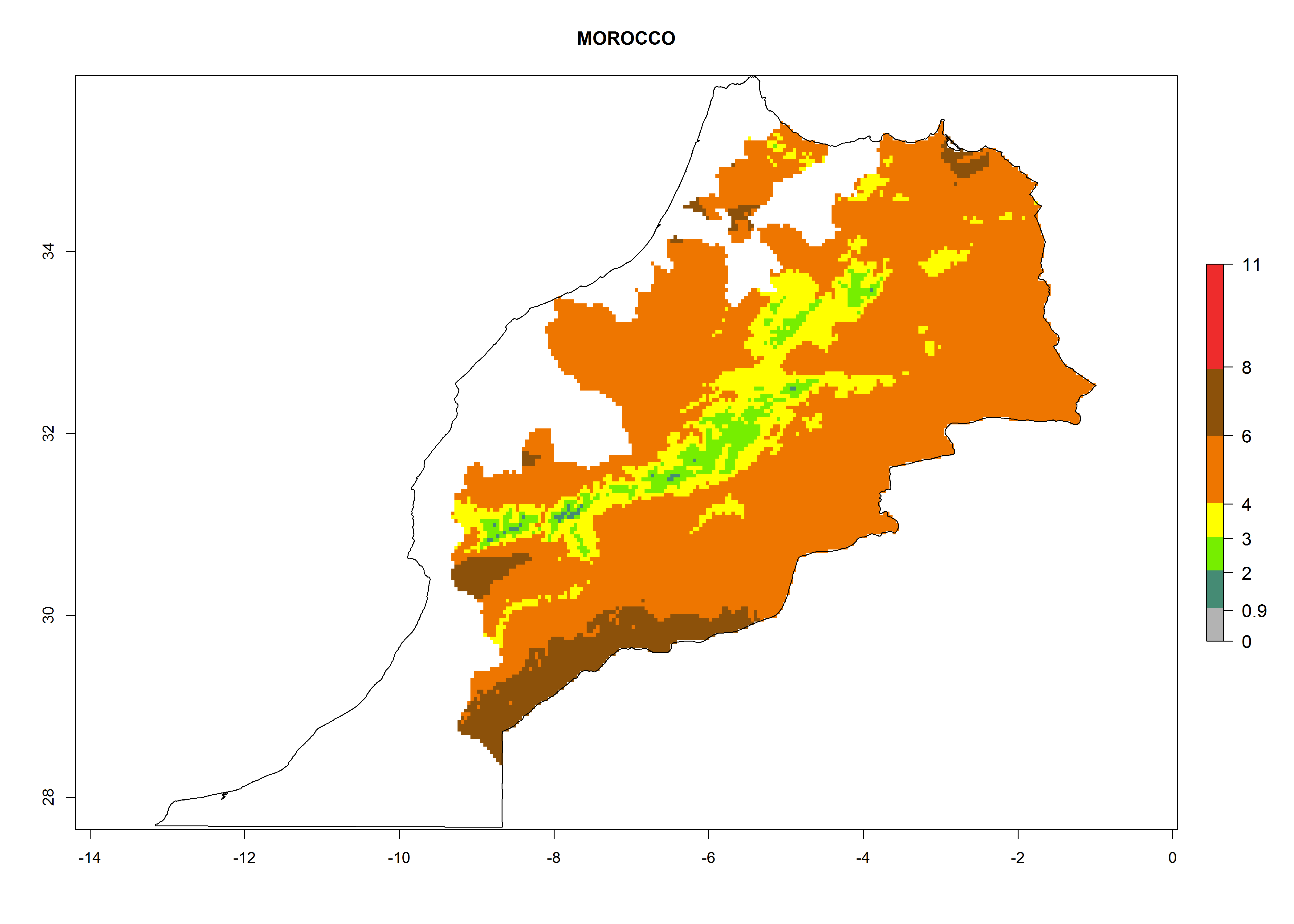 |
 |
| l) Mozambique
|
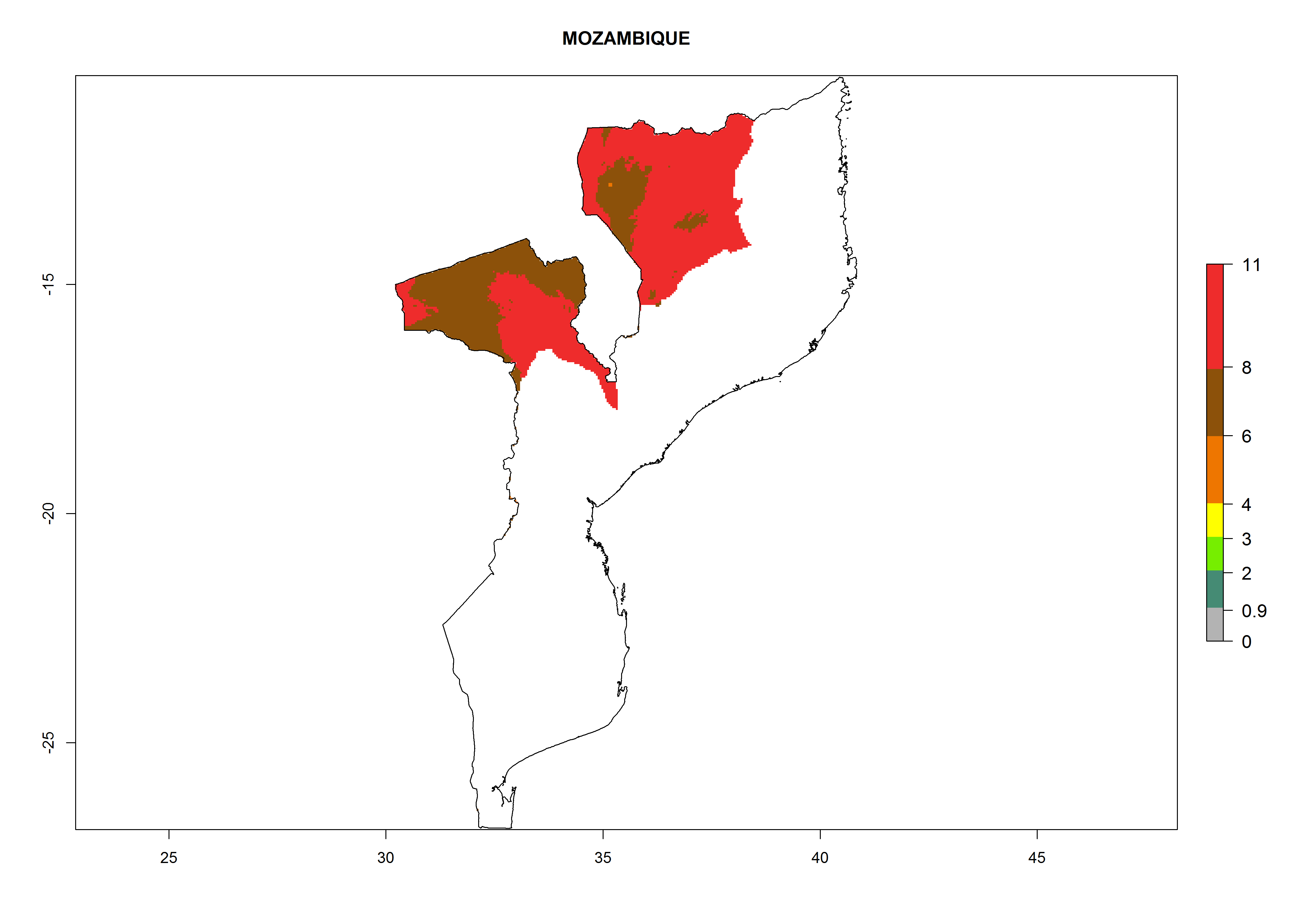 |
 |
| m) Rwanda
|
 |
|
| n) South Africa
|
 |
 |
| o) Tanzania
|
 |
 |
| p) Tunisia
|
 |
 |
| q) Uganda
|
 |
 |
| r) Zambia
|
 |
 |
| s) Zimbabwe
|
 |
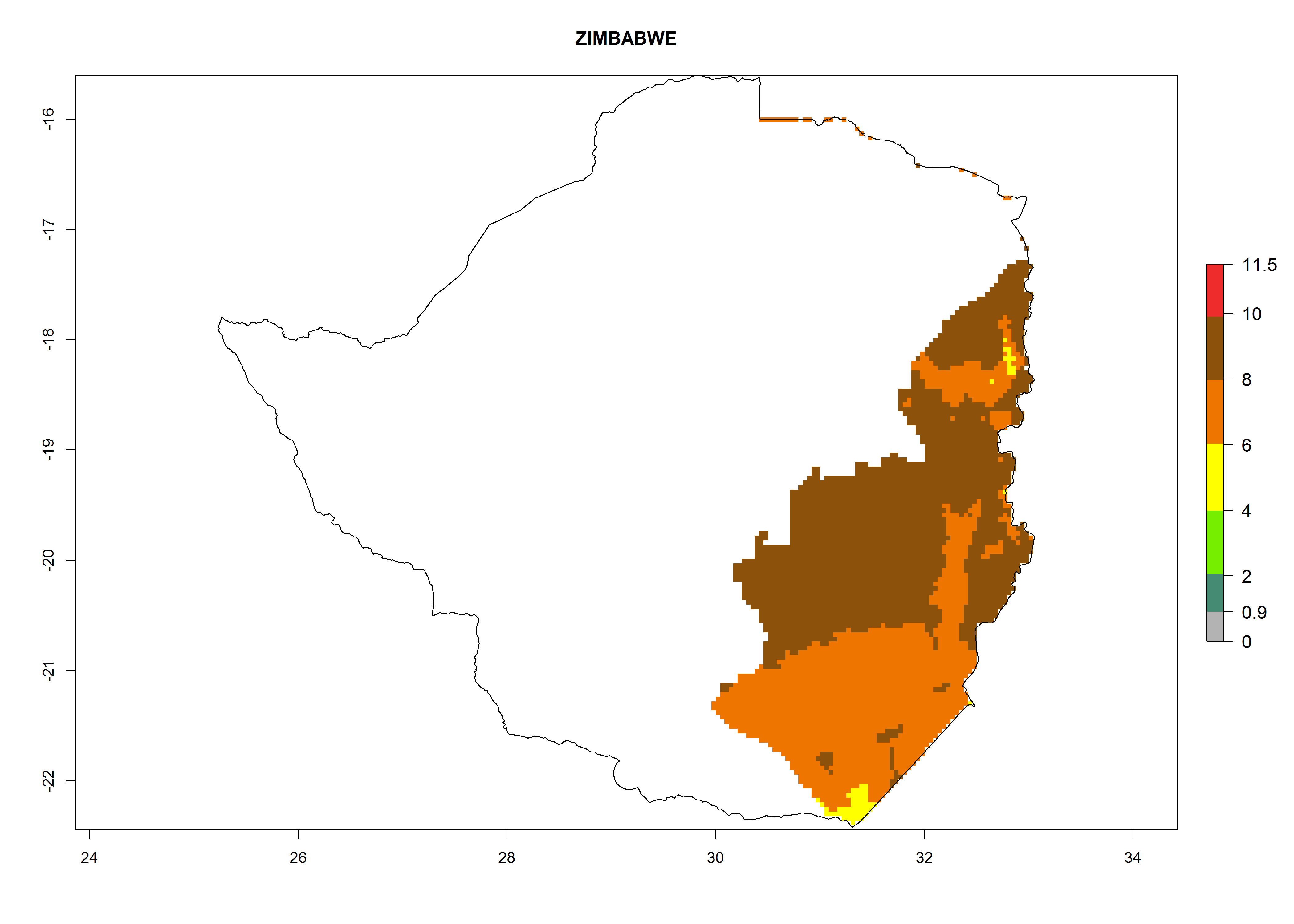 |
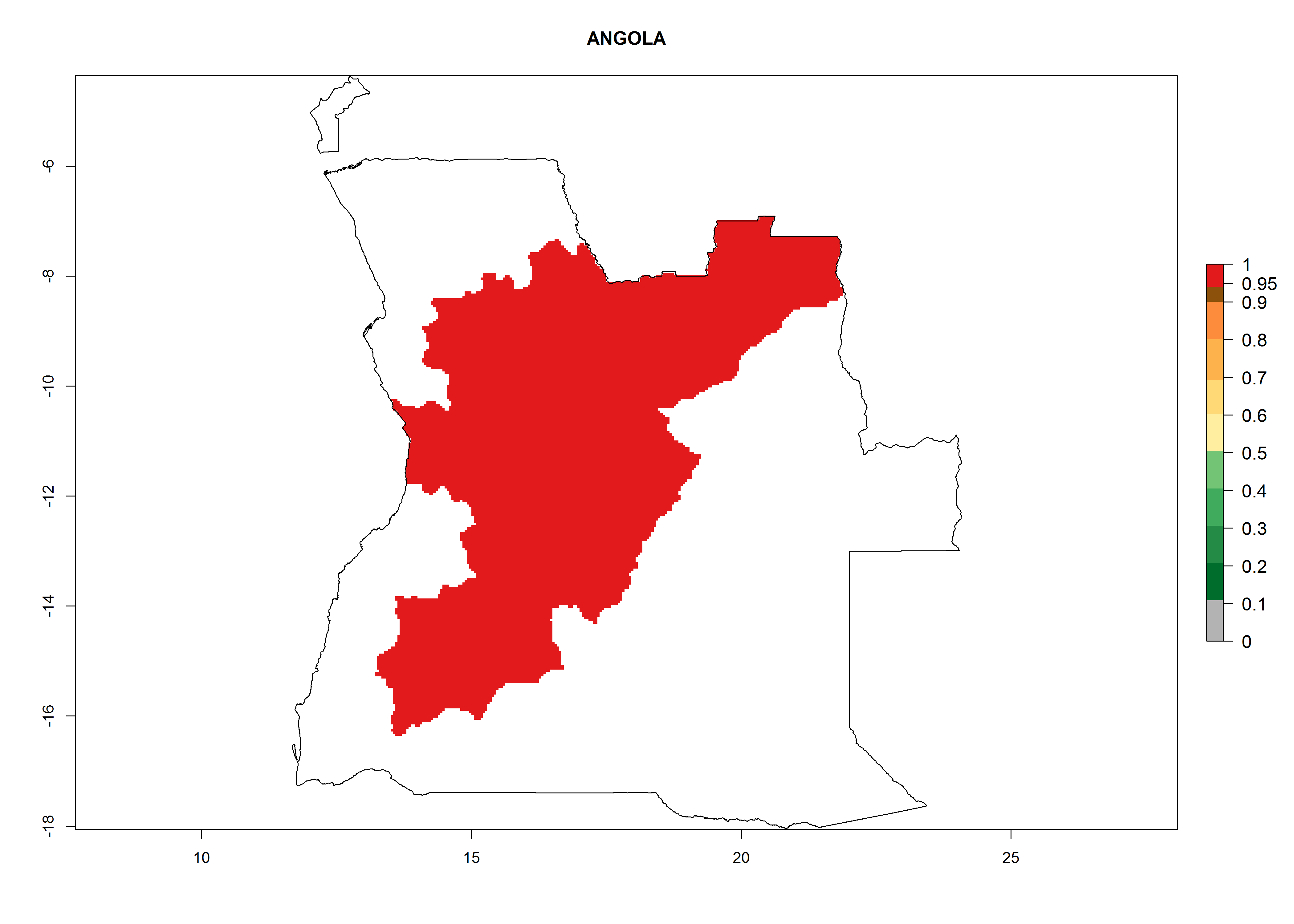
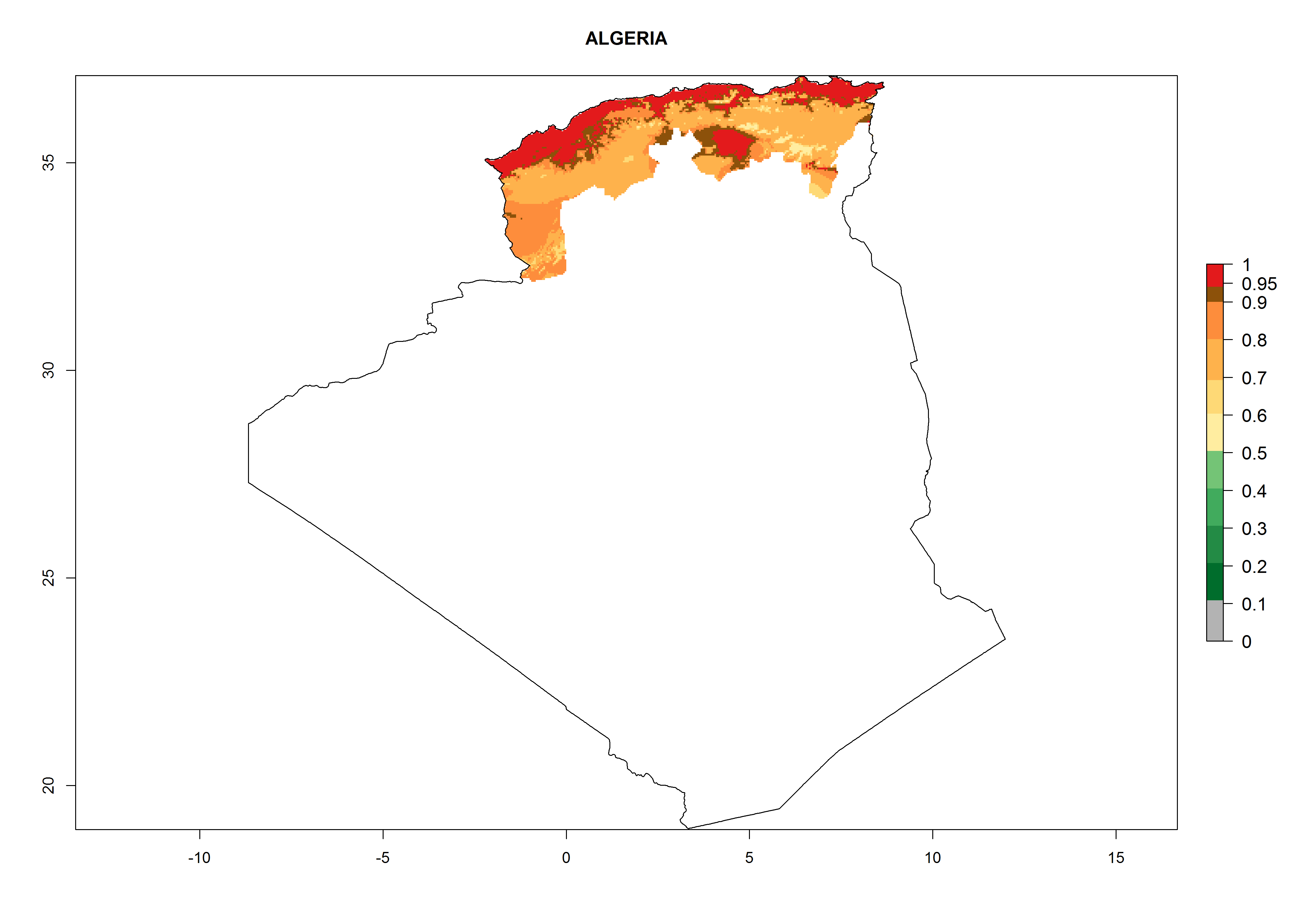
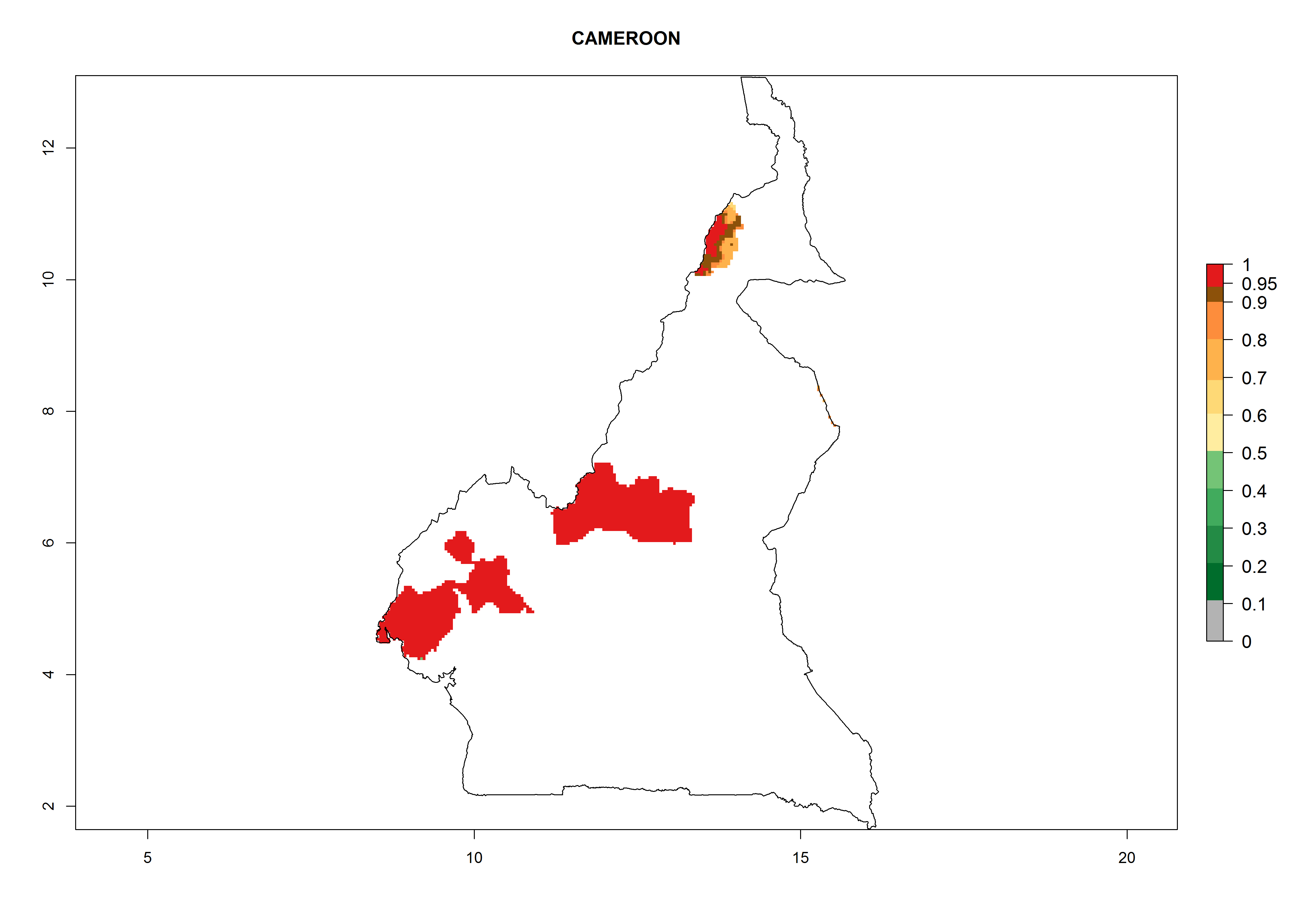
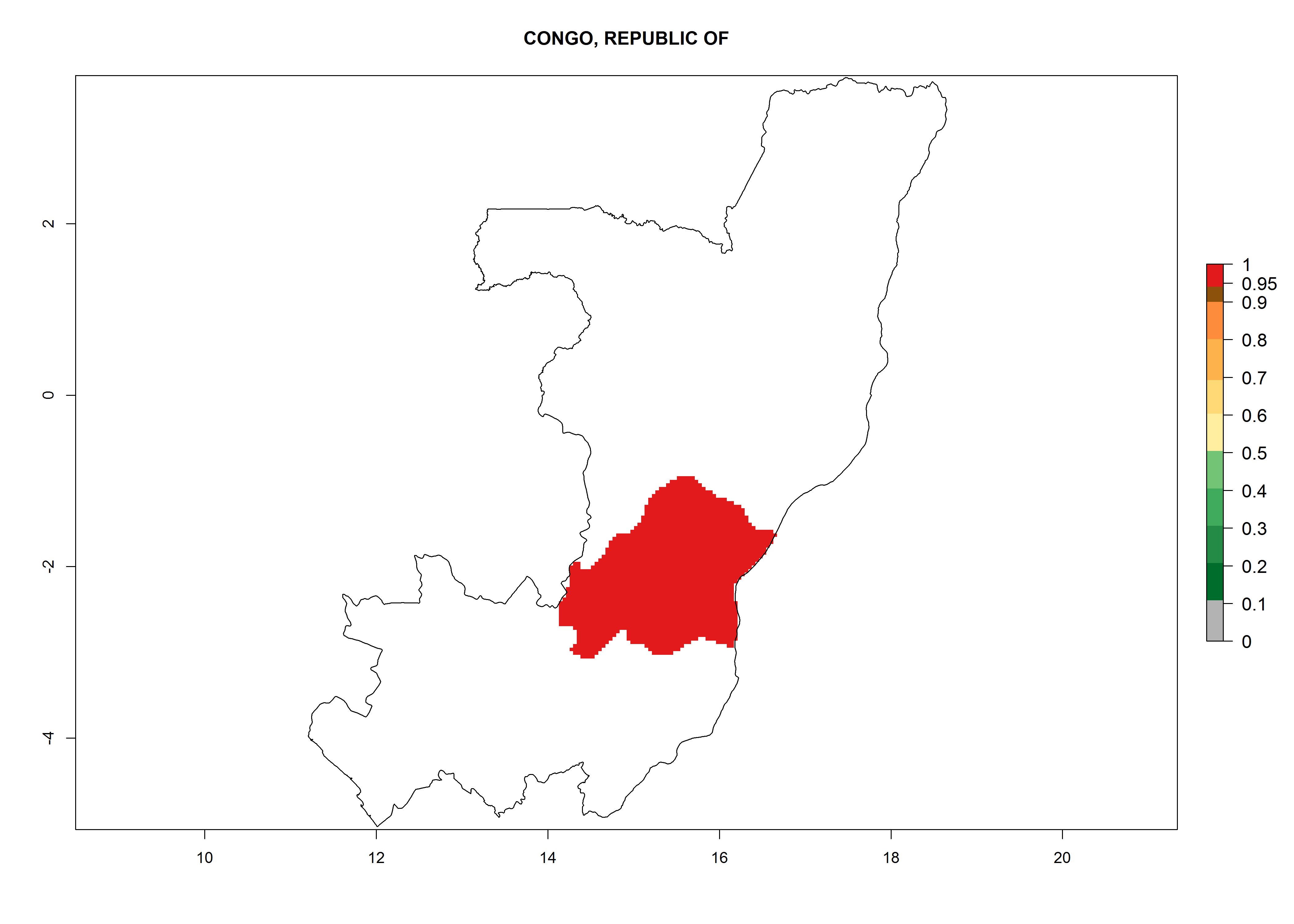
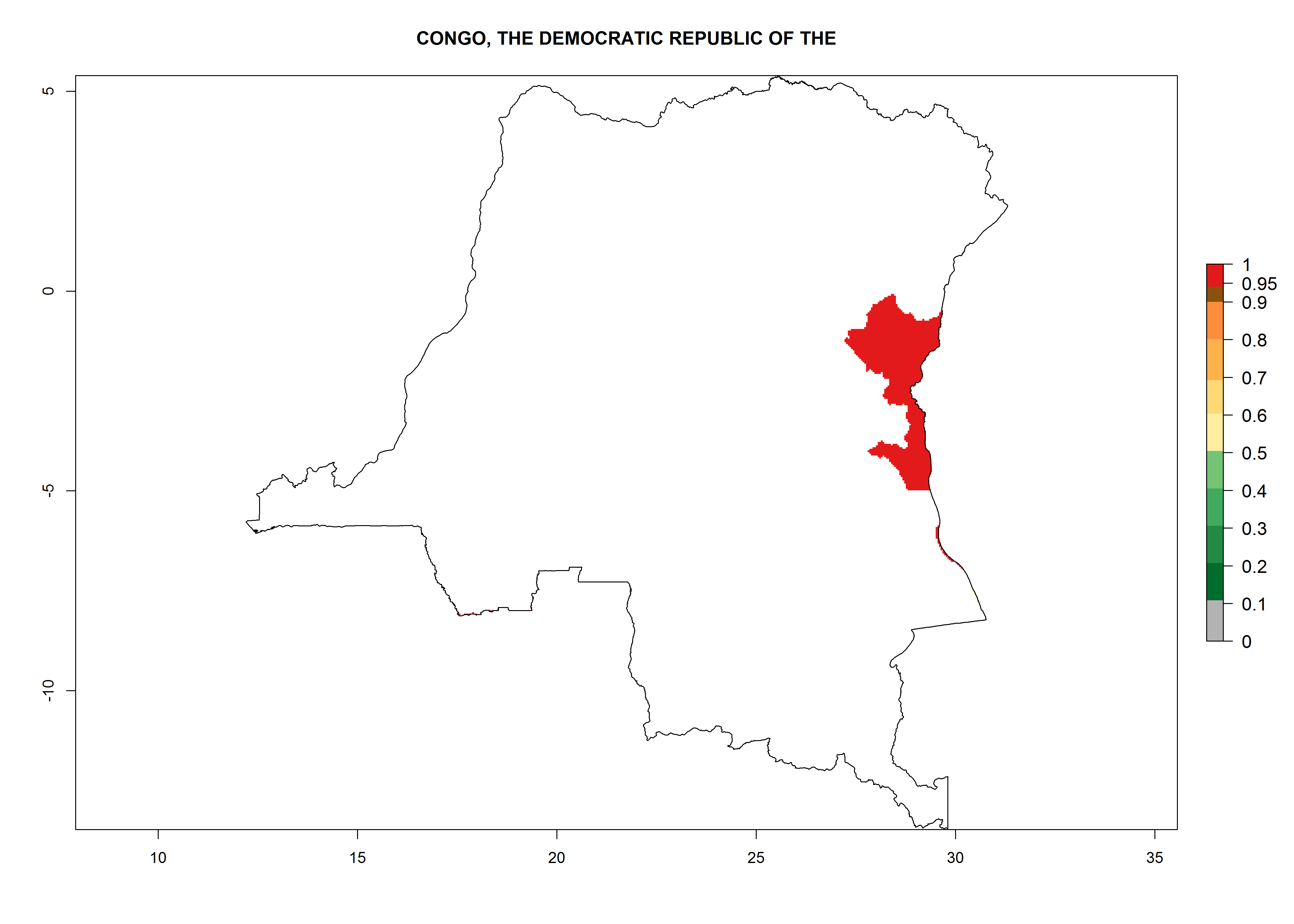
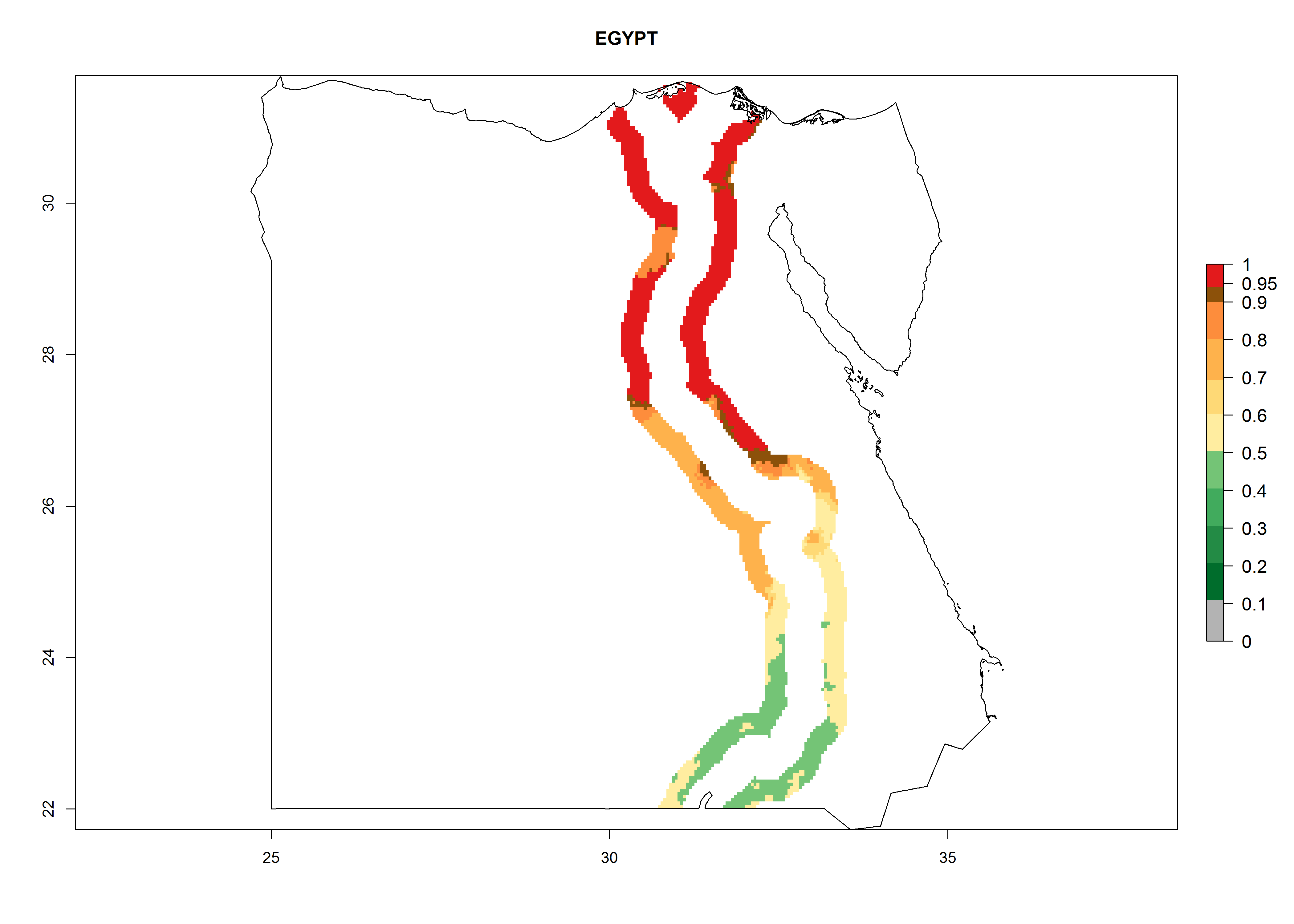
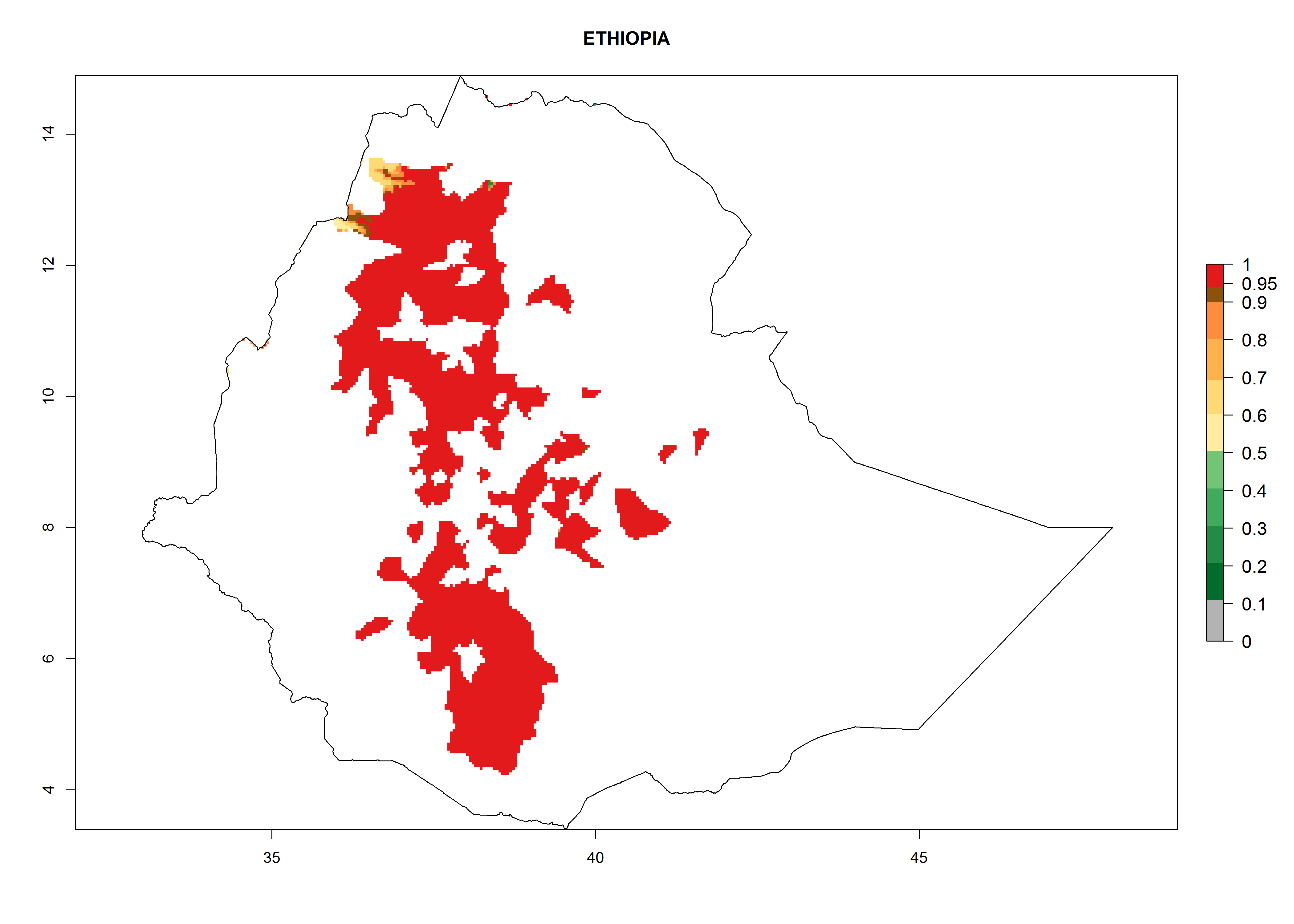
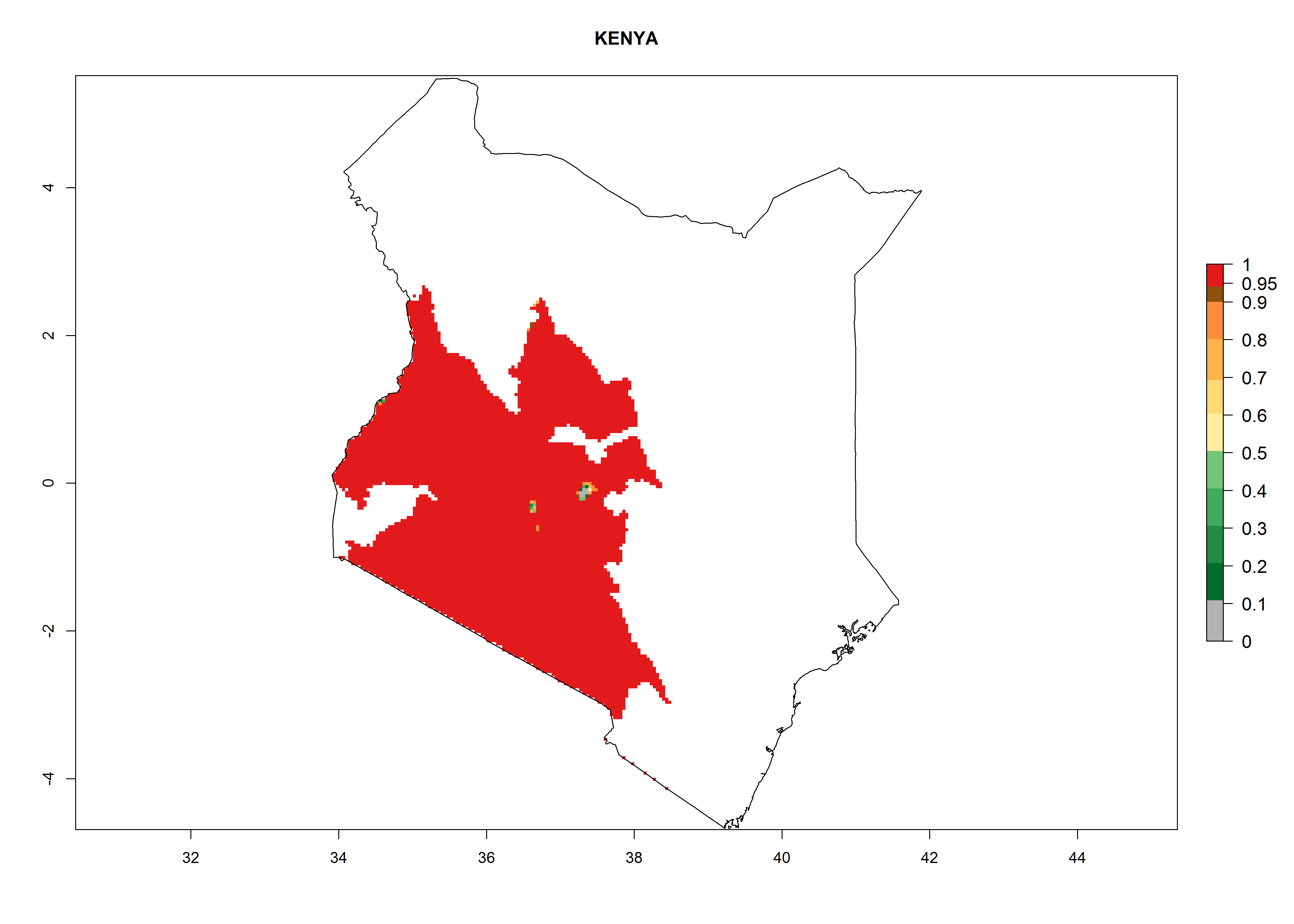
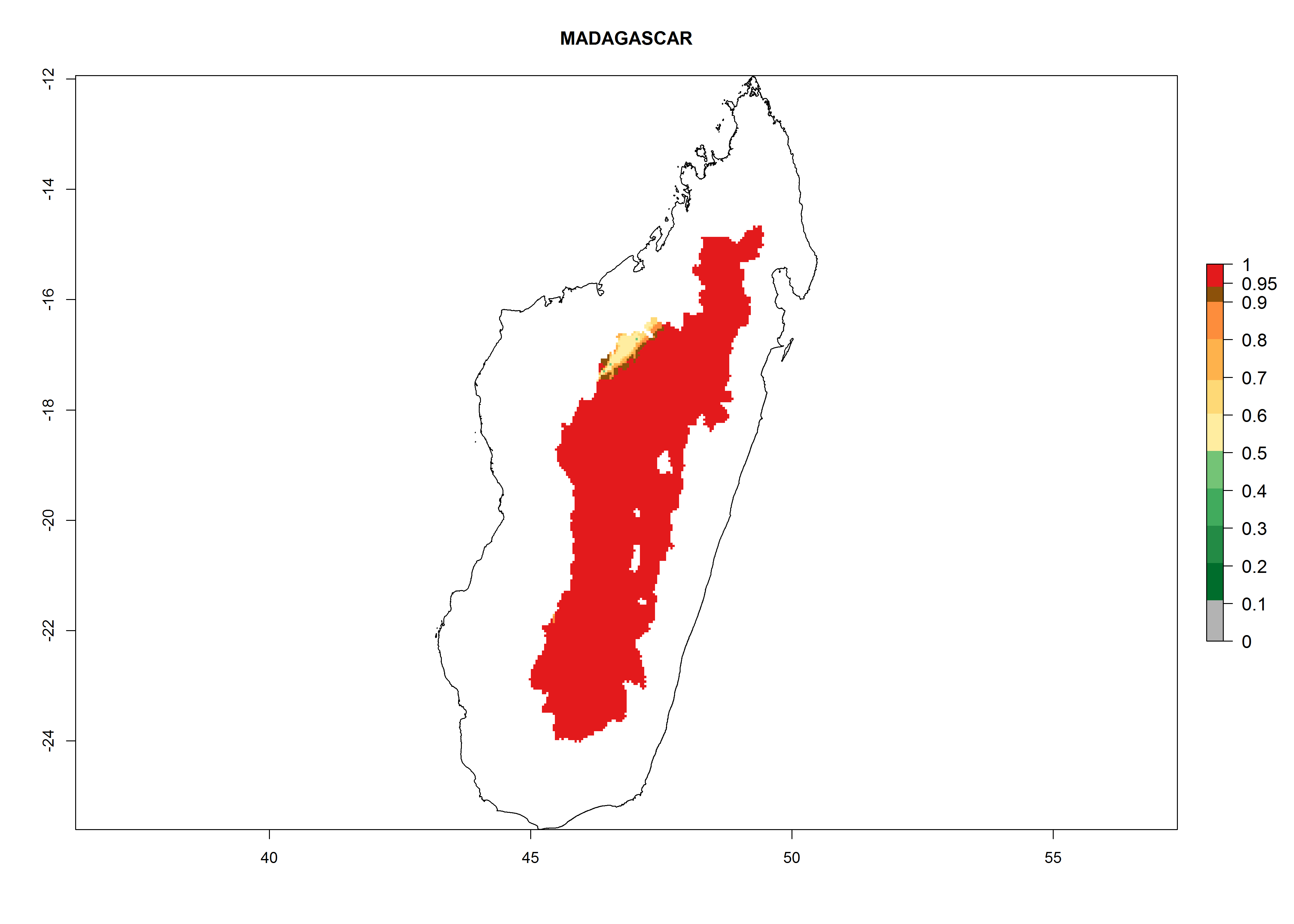
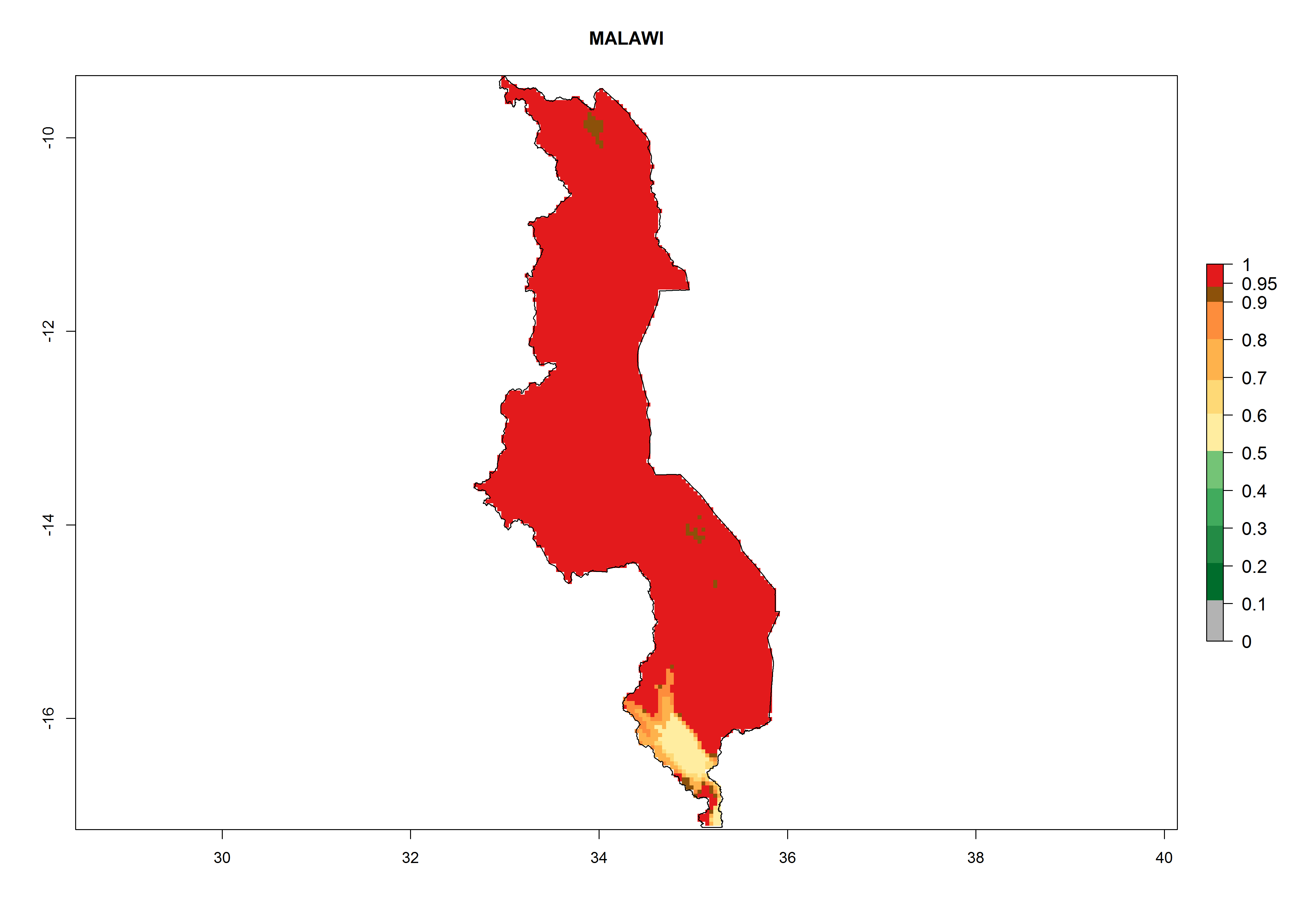


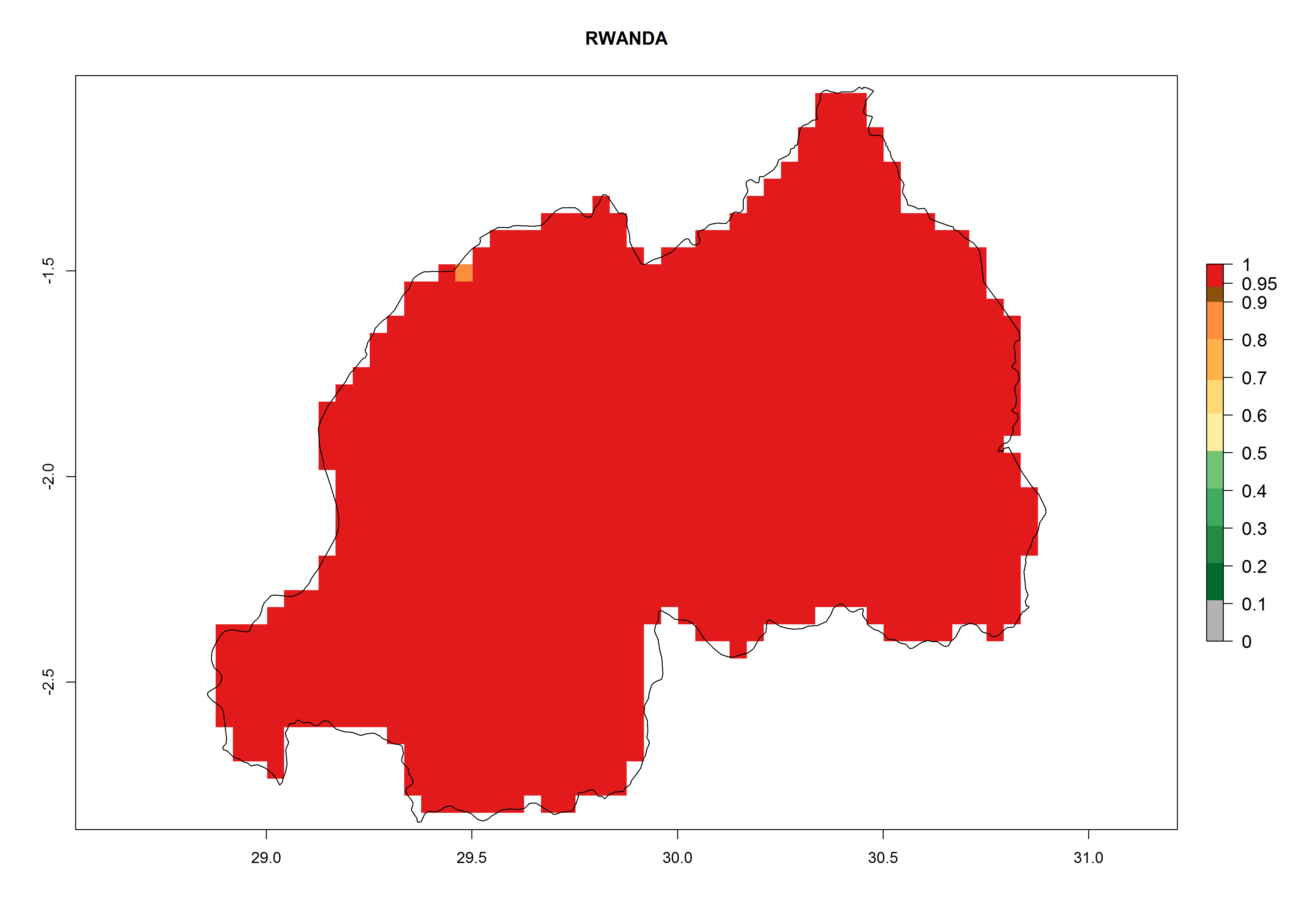
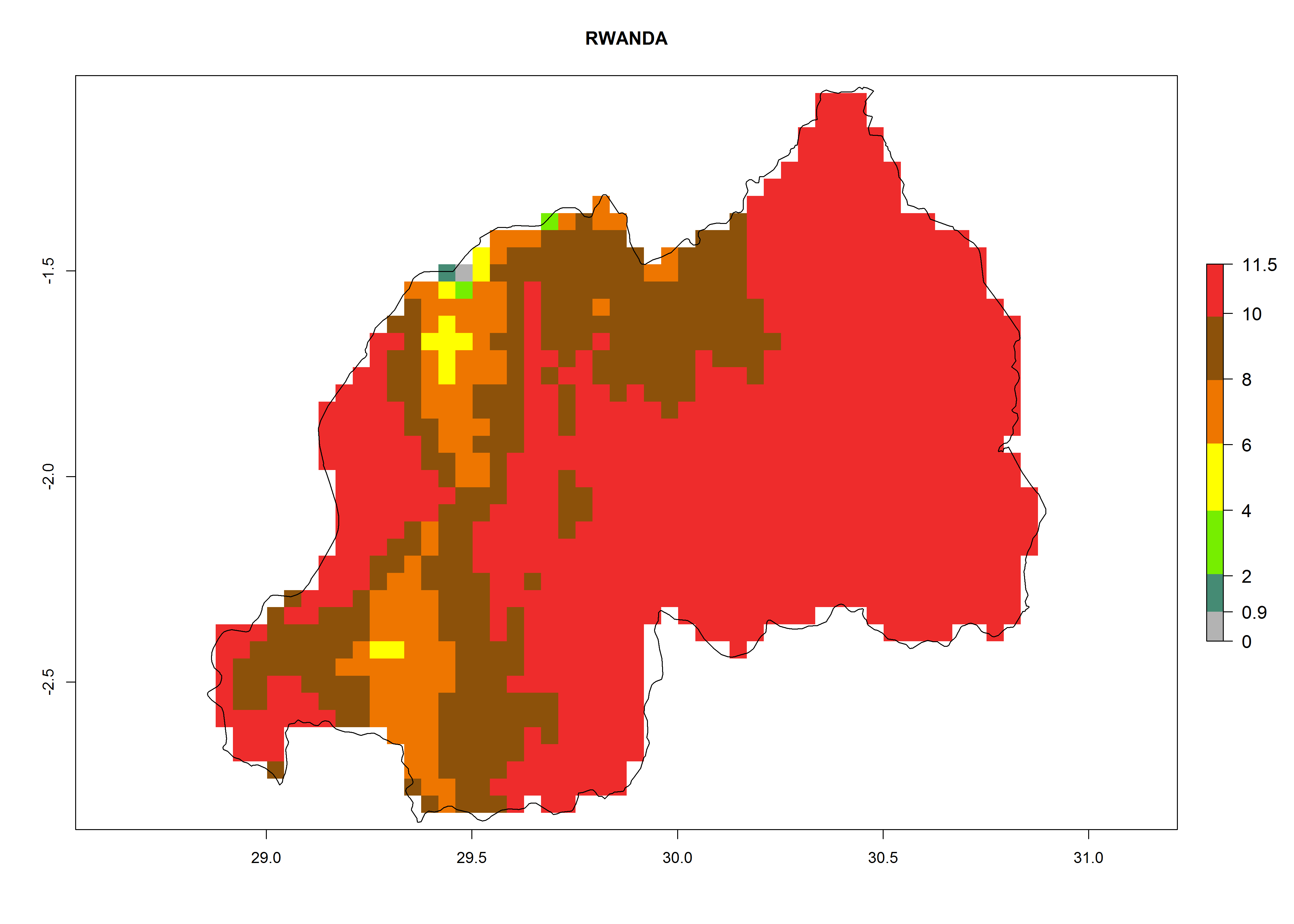
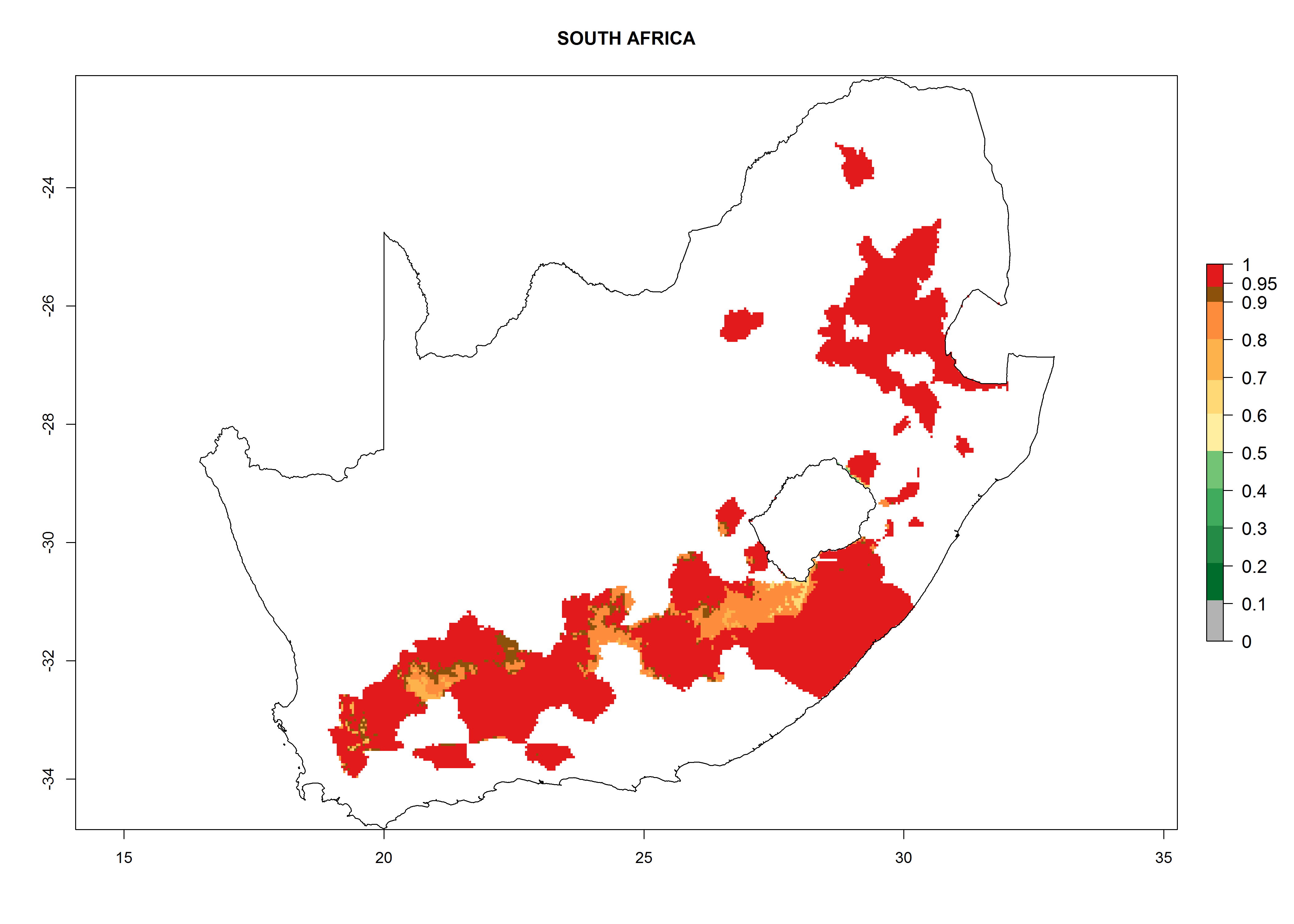
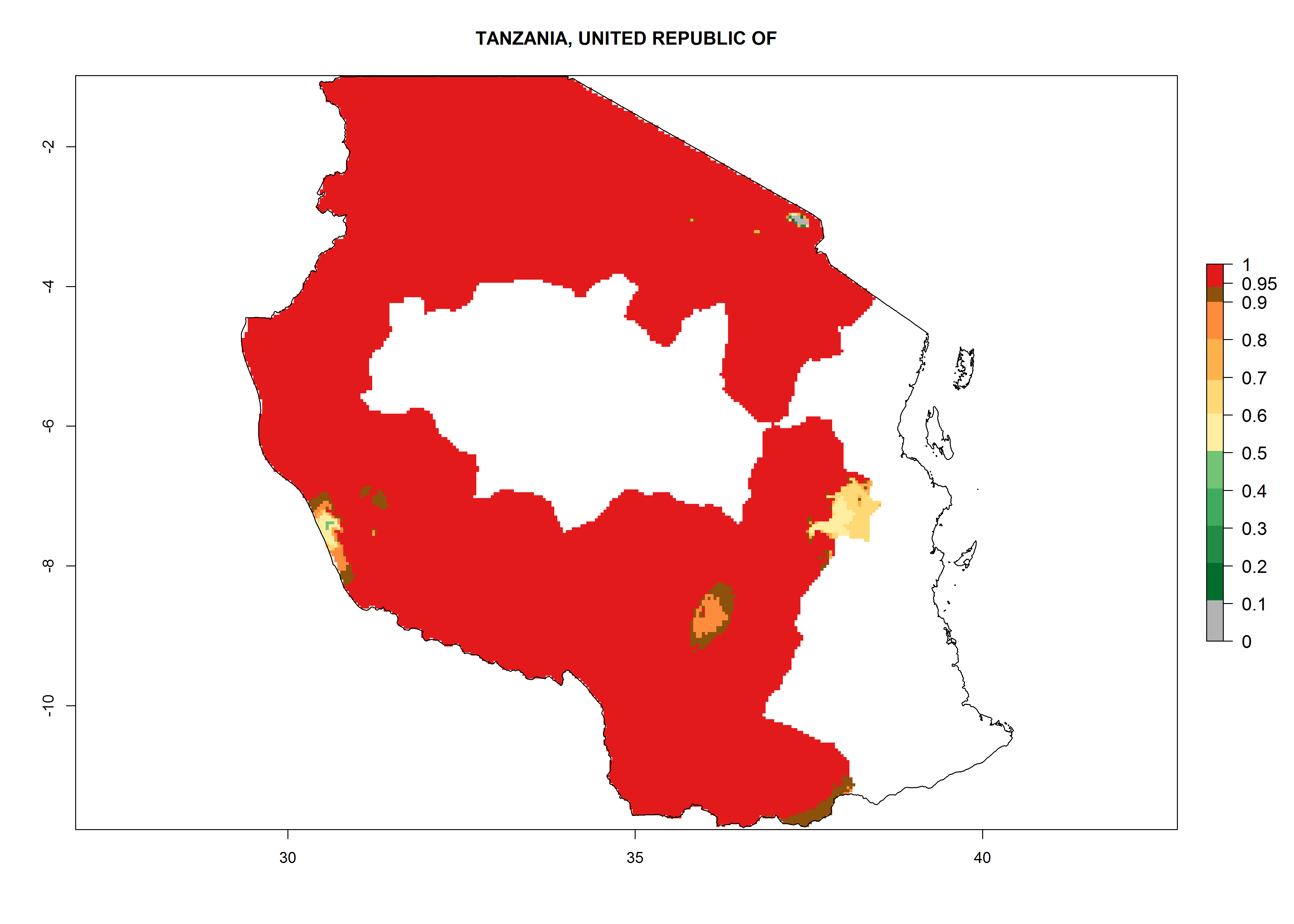
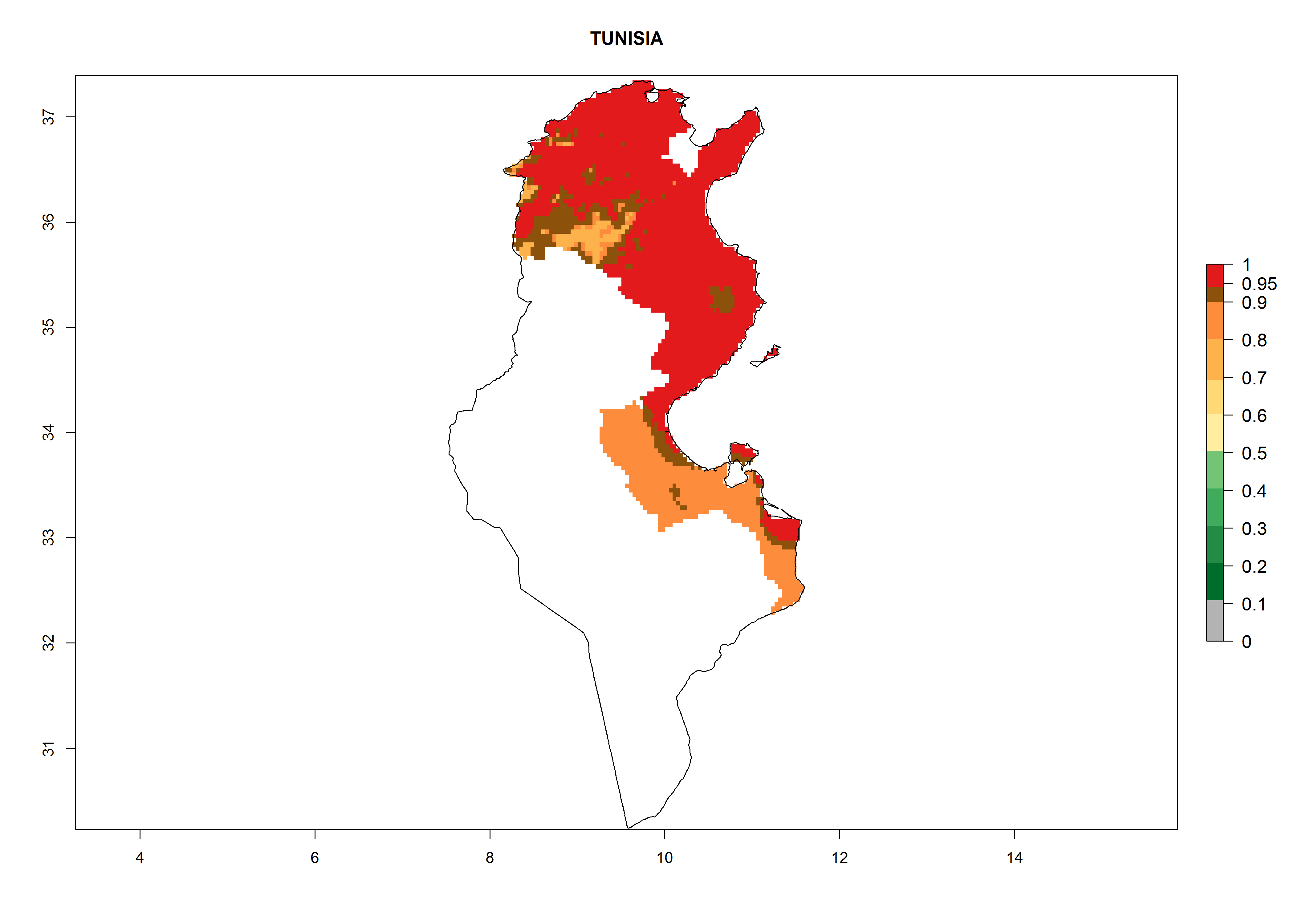

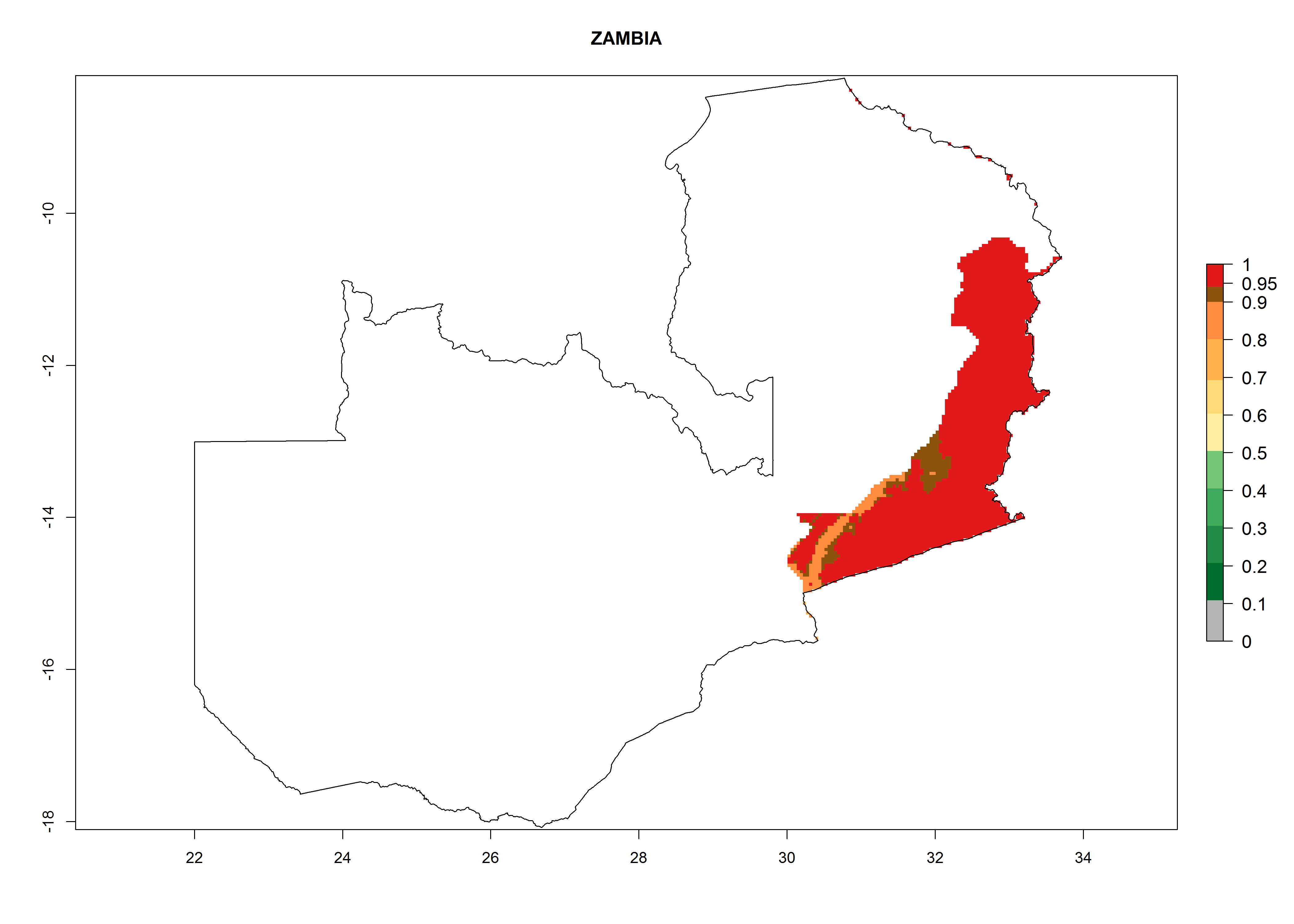
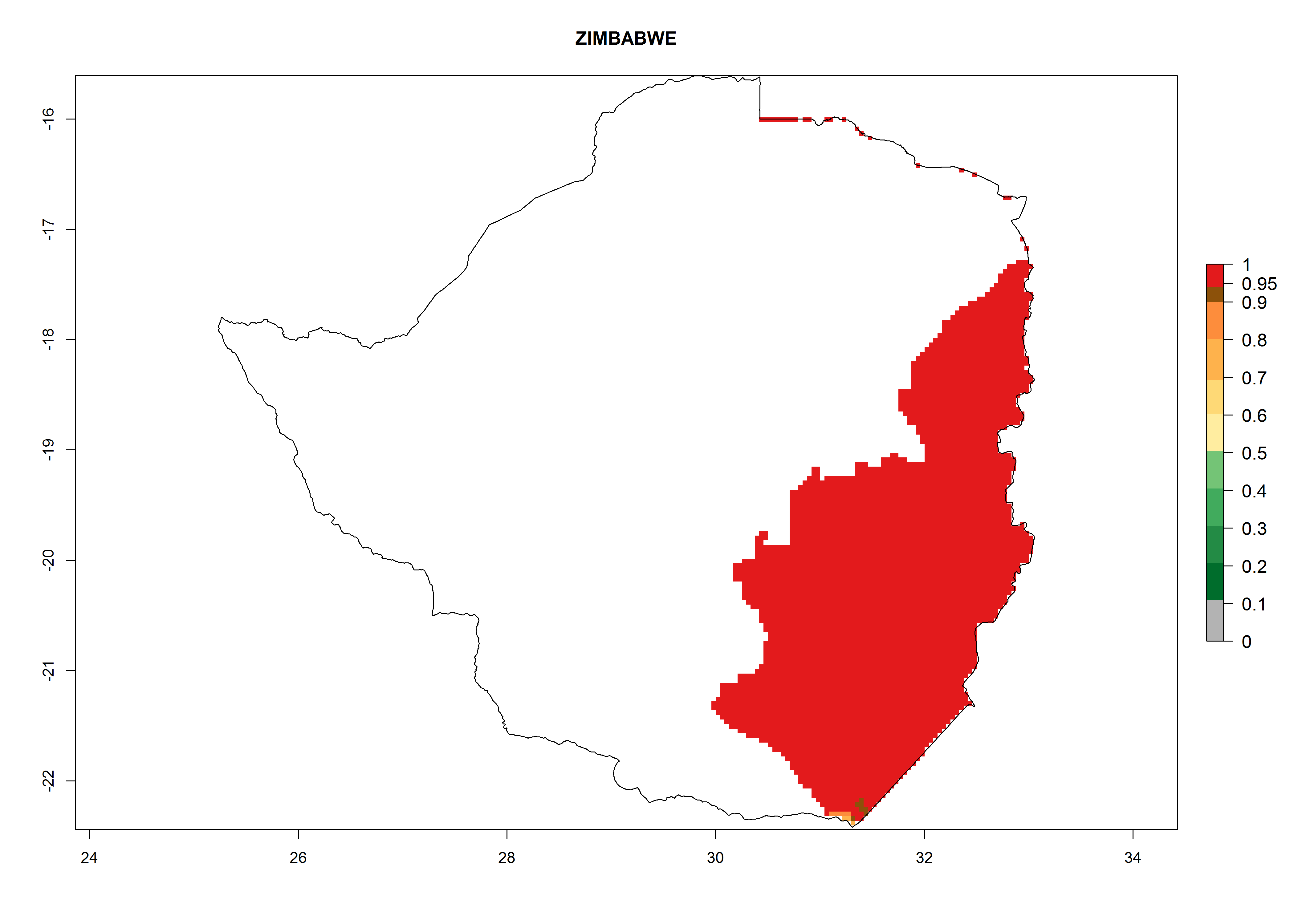
Figure 6. ERI, abundance (GI, potential damage), and activity (AI, potential population growth) of the Guatemalan potato tuber moth, Tecia solanivora, in potato production regions of selected African countries according to model predictions for the year 2000. An ERI>0.95 is associated with potential permanent establishment.
Phytosanitary measures
Infestation of potato tubers with eggs or young larvae of T. solanivora is hard to detect. Only a ban on importation of potato tubers from countries where the pest is present can prevent the dissemination effectively. EPPO recommends to avoid importation from countries with pest presence and to monitor the absence of the pest in countries that are still pest free. Furthermore, EPPO recommends importation of clean potatoes without soil and the use of new or disinfected packing materials (EPPO 2005). The national food safety authority of Peru carries out extensive surveys in which pheromone traps are installed and potato tubers in the Peruvian potato-growing zones are sampled.
Adaptation to risk avoidance at farm level
T. solanivora is difficult to control, as the larvae feed only inside potato tubers where they are hard to reach. Effective T. solanivora risk management therefore needs to focus on prevention, which is most effective when a range of protective methods is included.
Avoid infestation of pest-free potato-growing zones. Infested seed facilitates the fast spread of the pest and new establishment of the pest, if weather conditions are favorable. Pest-free seed tubers can be obtained from pest-free potato-growing areas. In the Andes these areas are located approx. above 3,500 masl, or where high precipitation impedes the establishment of T. solanivora.
Crop Management
Adequate hilling. In the field larvae need to enter the soil to reach potato tubers. The infestation of potato tubers decreases with the depth in soil where they develop. Therefore potato tubers need to be covered well with soil during the whole cultivation period.
Avoiding dry soil conditions. Dry weather conditions favor multiplication of T. solanivora in the field as the number of eggs laid decreases strongly with higher soil moisture; larvae drown in highly moist soil as well. The sowing date should be adjusted to avoid long periods of dry weather, especially in the weeks before harvest. If there is little rain, sprinkler irrigation can be used to moisten the soil.
Timely and complete harvest. High tuber damage at harvest has been frequently caused by delayed harvests, together with dry weather conditions that enable the development of several generations in the field. During dry periods, timely harvest is crucial to avoid tuber infestation. Early-maturing varieties can contribute to reduce the risk of infestation. Leftover tubers from harvest are frequently infested by T. solanivora and are a source of infestation for neighboring potato fields and following cropping periods. At harvest, the potato tubers need to be removed quickly from the field. All potato tubers need to be collected from the field, including those that are not appropriate for sale or consumption so that no habitat for T. solanivora larvae remains.
Monitoring with pheromone traps. Pheromone traps indicate the presence of T. solanivora in the field by attracting male adults. One trap should be placed in the center of the field and the number of males caught should be recorded weekly. Damage thresholds of 50–200 males/week are recommended for application of chemical control. However, control in the field is difficult and efficacy of chemical control is variable. The abovementioned measures like sprinkler irrigation, high hilling, or timely harvest might help to limit the damage.
Storage Management
Clean storage facilities. Infested potato stores are a source of contamination for tubers stored after harvest and in surrounding fields and stores. Before potato tubers are stored, the stores need to be cleaned thoroughly. Eggs, larvae, and pupae frequently found in cracks of the walls, the soil, or in bags or boxes used for storage need to be removed. Only healthy tubers should be selected for storage.
Biological control. Phthorimaea operculella granulovirus (PhopGV) products are regionally available and reduce infestation by 90–100% when they are dusted on potato tubers at the beginning of the storage period. Larvae get infected with granulovirus when they feed on the tubers and die at the end of their larval development. Furthermore, the abrasive effect of the powder (kaolin, calcium carbonate, or others), which is the basis of the formulation, also kills much of the young larvae. All potato tubers stored should be powdered with PhopGV previously.
Monitoring with pheromone traps. One pheromone trap should be installed per storage facility to detect the presence of T. solanivora early in order to take adequate control. Seriously infested batches of potato tubers need to be destroyed immediately.
Further reading
Castillo, G. 2005. Determinacion del ciclo de vida de las polillas de la papa Symmetrischema tangolias (Gyen) y Tecia solanivora (Povolny) bajo condiciones controladas de laboratorio. Undergraduate Thesis. Universidad Central del Ecuador, Quito, Ecuador. 144 pp.
CIP INIAP. 1997. Primer seminario taller internacional sobre manejo integrado de Tecia solanivora. Ibarra, Ecuador. CIP, INIAP, PNRT-Papa, FORTIPAPA. 61 pp.
CORPOICA. 2004. Memorias: III Taller Internacional sobre la polilla guatemalteca de la papa, Tecia solanivora. Cartagenas de Indias, Colombia. 16–17 de octubre 2003. Centro Internacional de la Papa (CIP), 2005. Lima, Peru, 137 pp.
Dangles, O., C. Carpio, A. Barragan, J. Zeddam, and J. Silvain. 2008. Temperature as a key driver of ecological sorting among invasive pest species in the tropical Andes. Ecological Applications 18(7): 1795–1809.
European Plant Protection Organisation (EPPO). 2005. Tecia solanivora. EPPO Bulletin 35: 399–401.
FAOSTAT. 2014. Food and Agriculture Organization of the United Nations—Statistic Division. Rome, Italy. Available from http://faostat.fao.org.
Kroschel, J., and B. Schaub. 2013. Biology and ecology of potato tuber moths as major pests of potato. In Insect pests of potato—Global perspectives on biology and management, ed. A. Alyokhin, C. Vincent, and P. Giordanengo, 165–192. Waltham, MA: Elsevier.
Niño, L. 2004. Revisión sobre la polilla de la papa Tecia solanivora en Centro y Suramerica. Suplemento de la Revista Latinoamericana de la papa. pp. 4–22.
Niño, L., and A. Notz. 2000. Patogenicidad de un virus granulosis de la polilla de la papa Tecia solanivora (Povolny) 1973 (Lepidoptera: Gelechiidae) en el estado Mérida. Venezuela. Boletín de Entomología Venezolana 15(1): 39–48.
Notz, A. 1995. Influencia de la temperatura sobre la biología de Tecia solanivora (Povolny) (Lepidoptera: Gelechiidae) criadas en tubérculos de papa Solanum tuberosum L. Boletín de Entomología Venezolana 11(1): 49–54.
Pollet, A., A. Barragán, and P. Iturralde. 2004. Conozca y maneje la polilla de la papa. Serie de divulgación: 3. Centro de biodiversidad y ambiente, escuela de biología. Pontificia Universidad Católica del Ecuador. 52 p.
Pollet, A., A. Barragán, J.L. Zeddam, and X. Lery. 2003. Tecia solanivora, a serious biological invasion of potato cultures in South America. International Pest Control 45(3): 139–144.
Povolny, D. 1973. Scobipalpopsis solanivora sp. n. A new pest of potato (Solanum tuberosum) from Central America. Acta Universitatis Agriculturae 21: 133–145.
PUCE. 2004. Memorias II Taller Internacional de Polilla Guatemalteca: avances en investigación y manejo integrado de la polilla guatemalteca de la papa Tecia solanivora. Pontificia Universidad Católica del Ecuador, 2004. 216p.
Sporleder, M., and J. Kroschel. 2008. The potato tuber moth granulovirus (PoGV): use, limitations and possibilities for field applications. In Integrated pest management for the potato tuber moth, Phthorimaea operculella (Zeller)—A potato pest of global importance, ed. J. Kroschel and L. Lacey, 49–71, Tropical Agriculture 20, Advances in Crop Research 10. Weikersheim, Germany: Margraf Publishers.
Torres, F., A. Notz, and L. Valencia. 1997. Ciclo de vida y otros aspectos de biología de la papa Tecia solanivora Povolny (Lepidoptera: Gelechiidae) en el estado Táchira, Venezuela. Boletín de Entomología Venezolana 12(1): 81–94.
Villamizar, L., C. Espinel, E. Grijalba, J. Gómez, A. Cotes, L. Torres, G. Barrera, X. Lery, and J-L Zeddam. 2008. Isolation, identification and biocontrol activity of Colombian isolates of granulovirus from Tecia solanivora larvae. Insect Pathogens and Insect Parasitic Nematodes IOBC/WPRS Bulletin 31: 85–88.